Ren Longhui, Huang Wei, Cannon Ethalinda K S, Bertioli David J, Cannon Steven B
Interdepartmental Genetics Graduate Program, Iowa State University, Ames, IA, United States.
Department of Agronomy, Iowa State University, Ames, IA, United States.
Front Genet. 2018 Oct 9;9:454. doi: 10.3389/fgene.2018.00454. eCollection 2018.
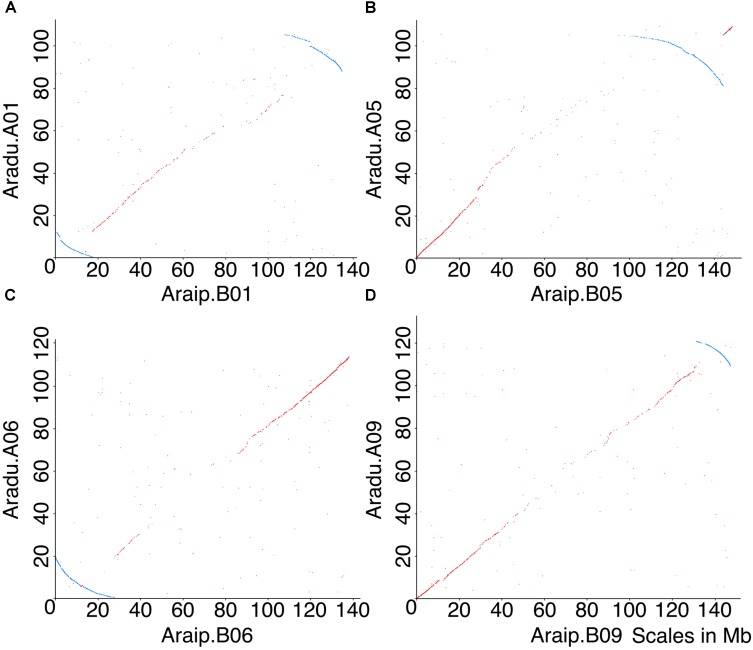
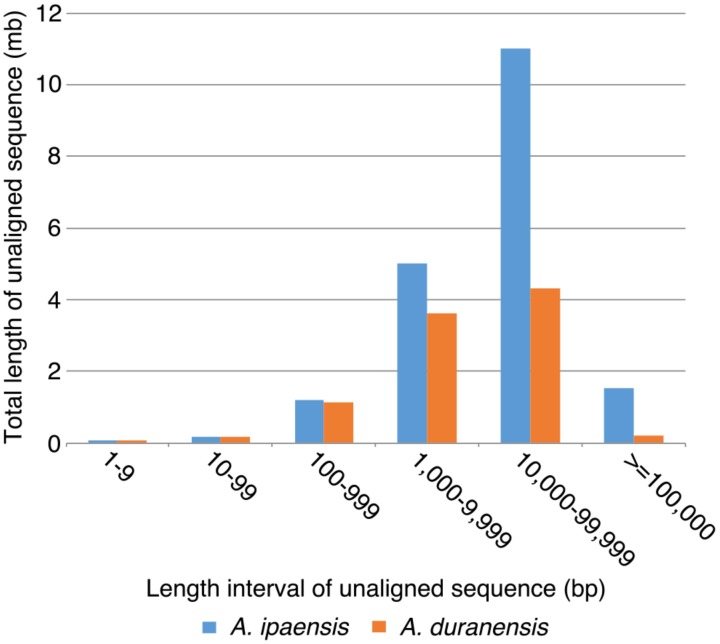
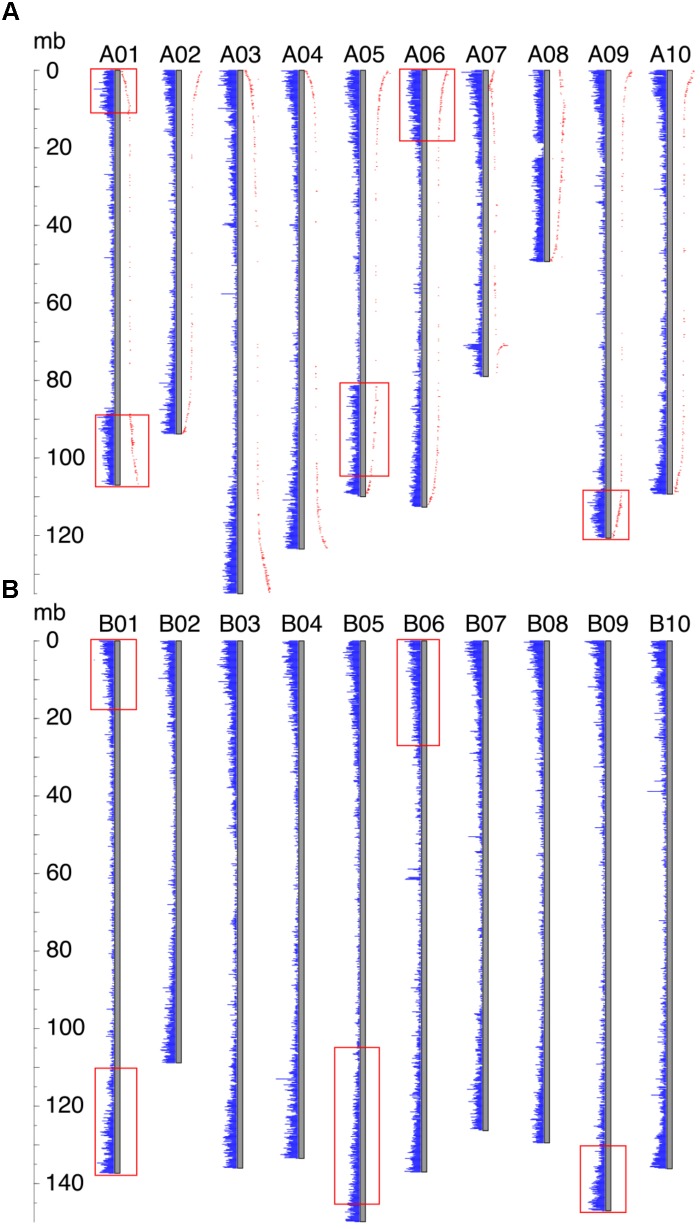
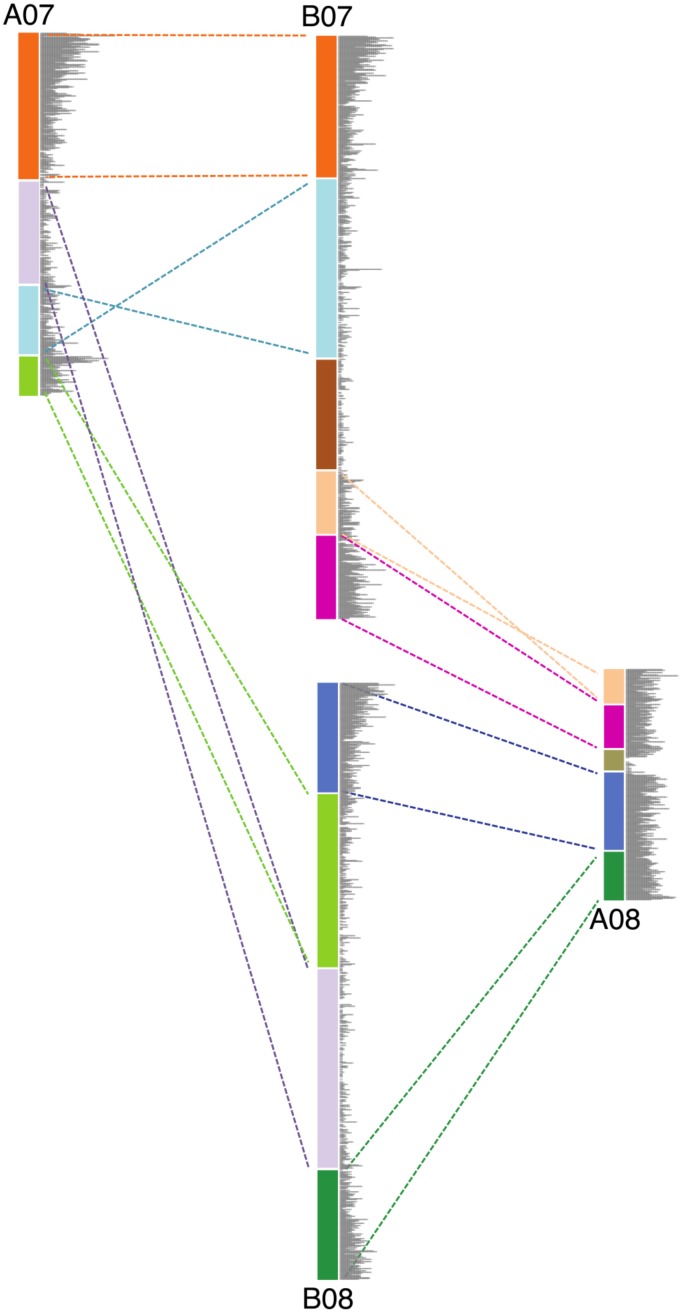
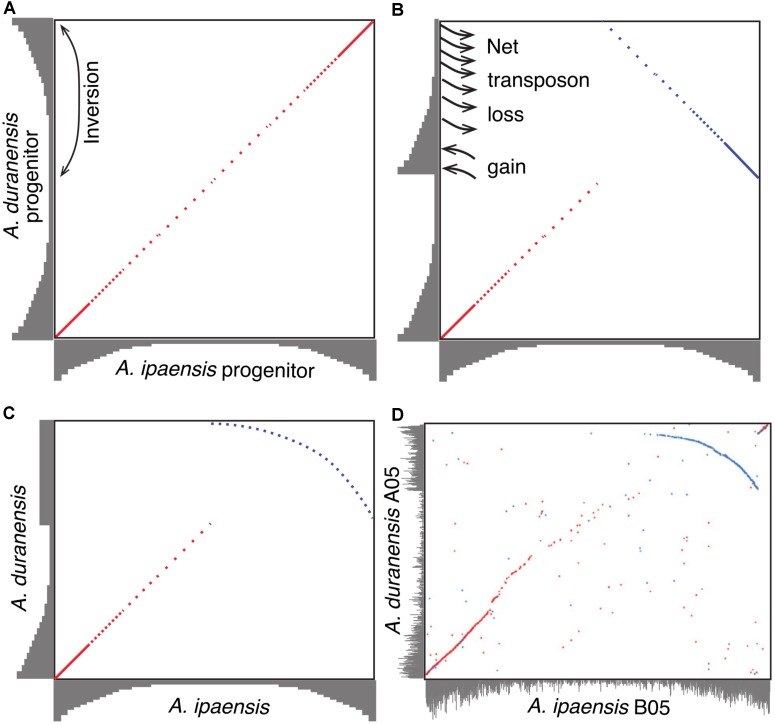
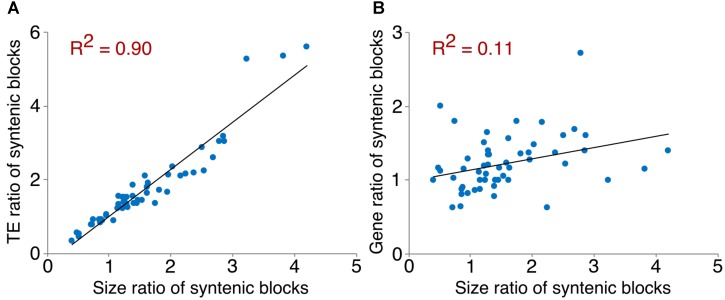
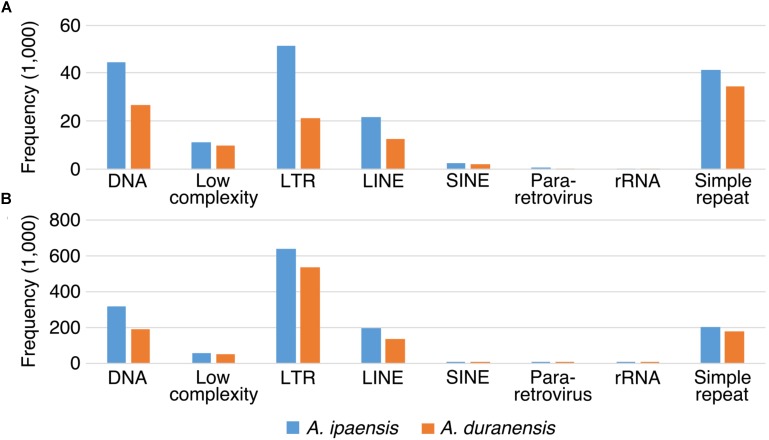
The factors behind genome size evolution have been of great interest, considering that eukaryotic genomes vary in size by more than three orders of magnitude. Using a model of two wild peanut relatives, and , in which one genome experienced large rearrangements, we find that the main determinant in genome size reduction is a set of inversions that occurred in , and subsequent net sequence removal in the inverted regions. We observe a general pattern in which sequence is lost more rapidly at newly distal (telomeric) regions than it is gained at newly proximal (pericentromeric) regions - resulting in net sequence loss in the inverted regions. The major driver of this process is recombination, determined by the chromosomal location. Any type of genomic rearrangement that exposes proximal regions to higher recombination rates can cause genome size reduction by this mechanism. In comparisons between and , we find that the inversions all occurred in . Sequence loss in those regions was primarily due to removal of transposable elements. Illegitimate recombination is likely the major mechanism responsible for the sequence removal, rather than unequal intrastrand recombination. We also measure the relative rate of genome size reduction in these two diploids. We also test our model in other plant species and find that it applies in all cases examined, suggesting our model is widely applicable.
考虑到真核生物基因组大小相差三个数量级以上,基因组大小进化背后的因素一直备受关注。利用两个野生花生近缘种A. duranensis和A. ipaensis的模型,其中一个基因组经历了大规模重排,我们发现基因组大小缩减的主要决定因素是发生在A. ipaensis中的一组倒位,以及随后倒位区域的净序列去除。我们观察到一种普遍模式,即新的远端(端粒)区域的序列丢失比新的近端(着丝粒周围)区域的序列获得更快,导致倒位区域的净序列丢失。这一过程的主要驱动因素是重组,由染色体位置决定。任何使近端区域暴露于更高重组率的基因组重排类型都可通过这种机制导致基因组大小缩减。在A. duranensis和A. ipaensis的比较中,我们发现倒位均发生在A. ipaensis中。这些区域的序列丢失主要是由于转座元件的去除。非法重组可能是负责序列去除的主要机制,而不是不等的链内重组。我们还测量了这两个二倍体中基因组大小缩减的相对速率。我们还在其他植物物种中测试了我们的模型,发现它适用于所有检测的案例,表明我们的模型具有广泛的适用性。