Department of Biological Sciences, University of Maryland, Baltimore, MD, USA.
Department of Biological Sciences, University of Maryland, Baltimore, MD, USA
Life Sci Alliance. 2019 Mar 5;2(2). doi: 10.26508/lsa.201800150. Print 2019 Apr.
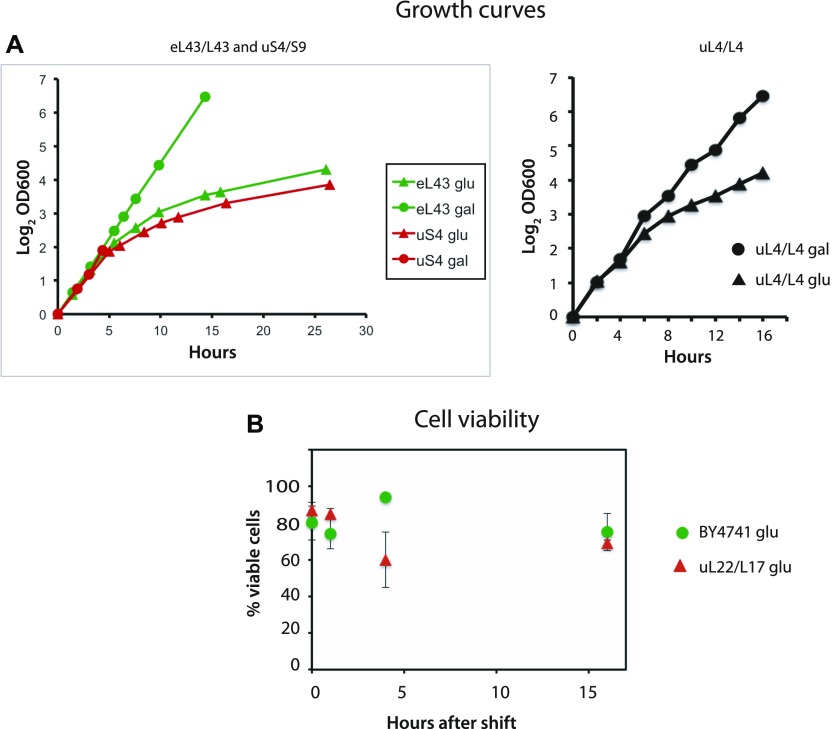
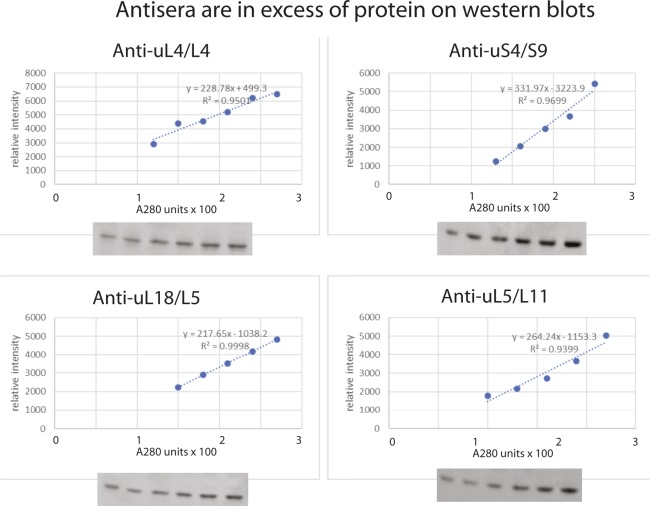
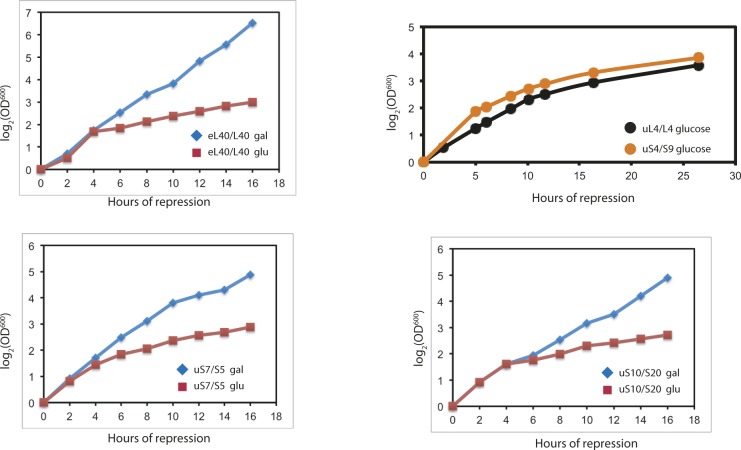
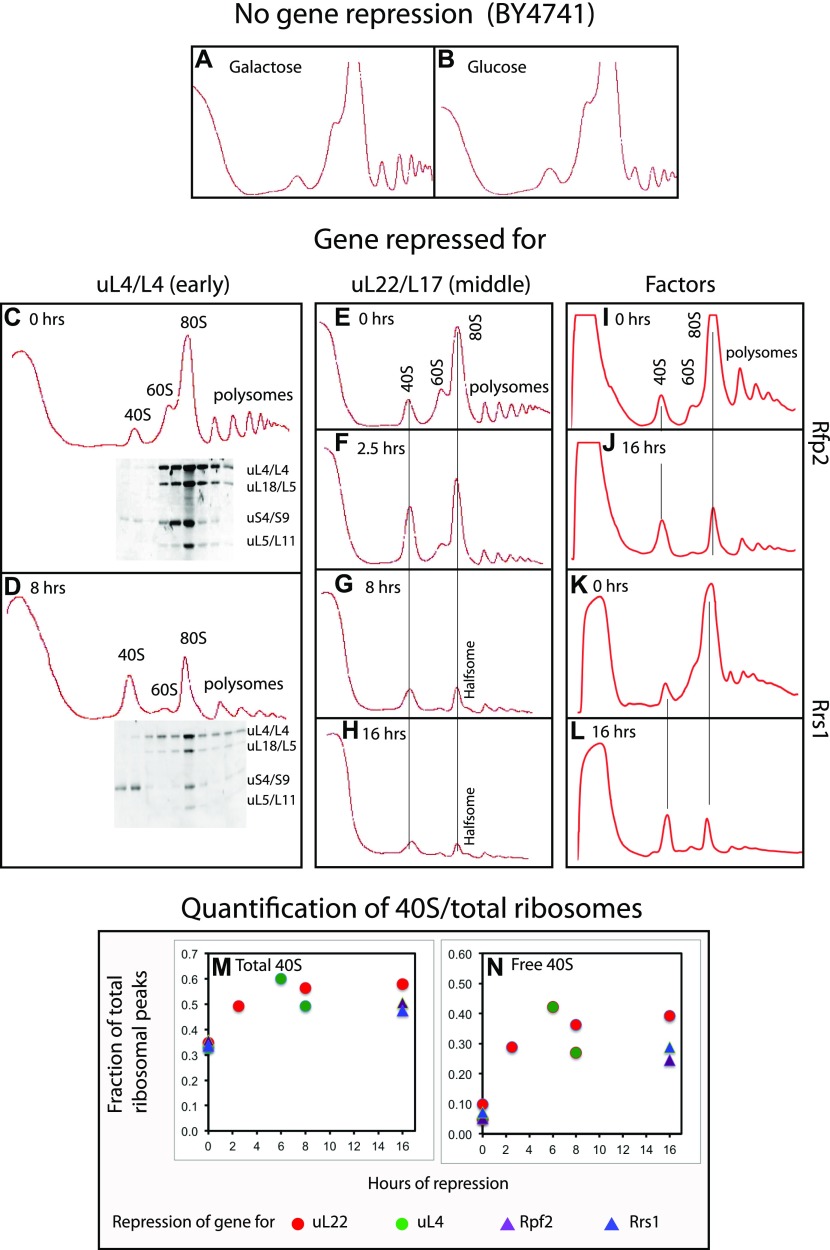
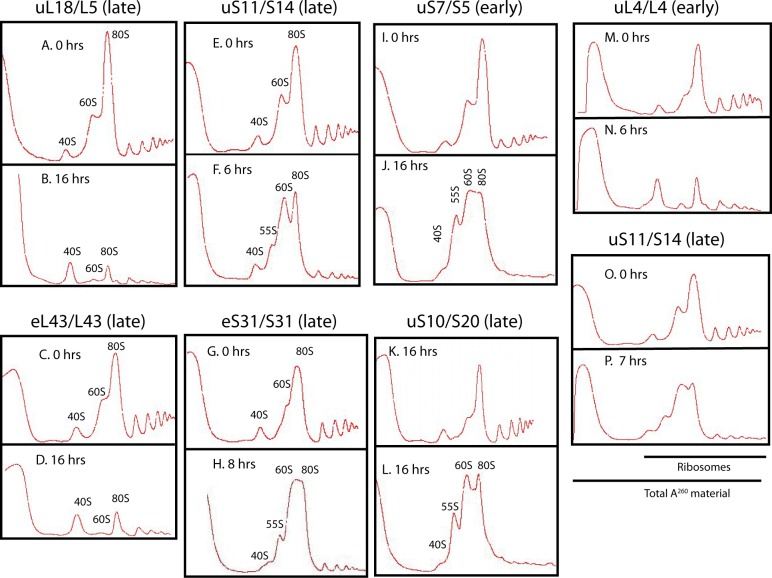
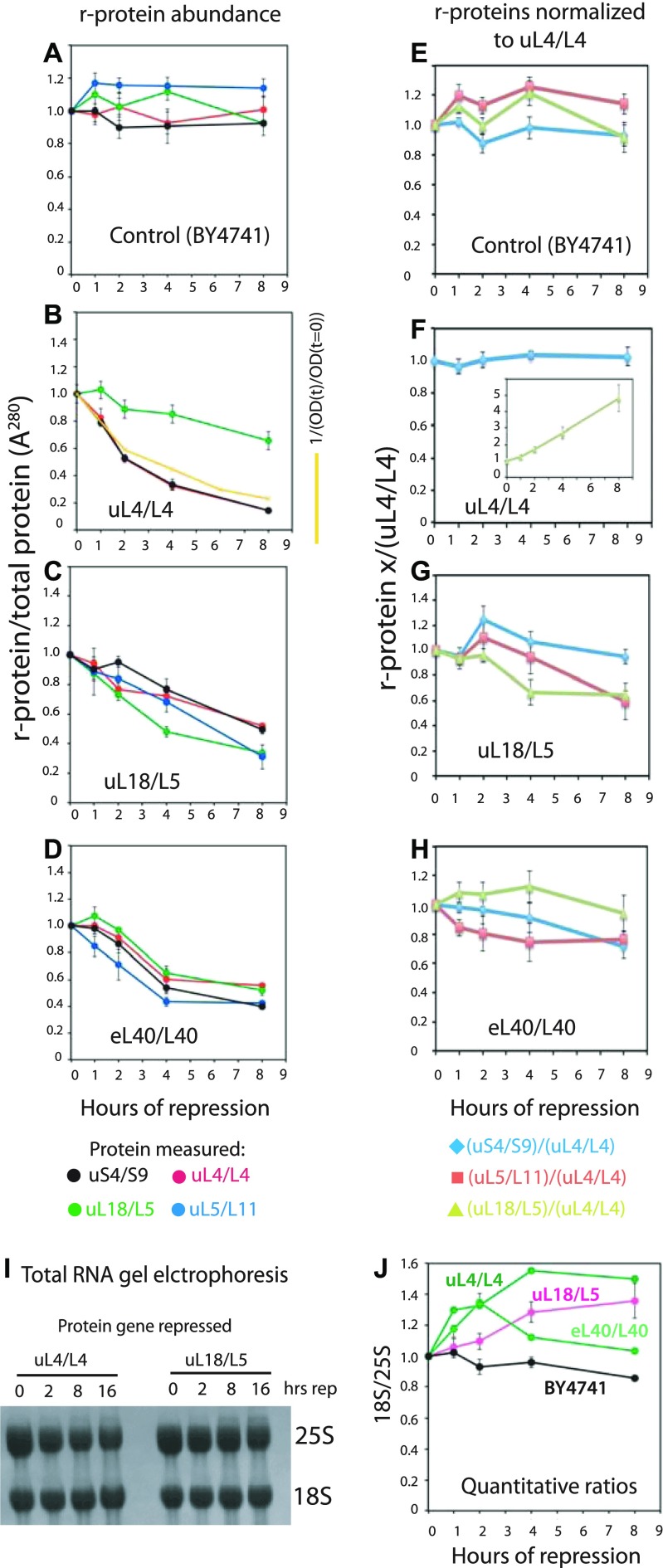
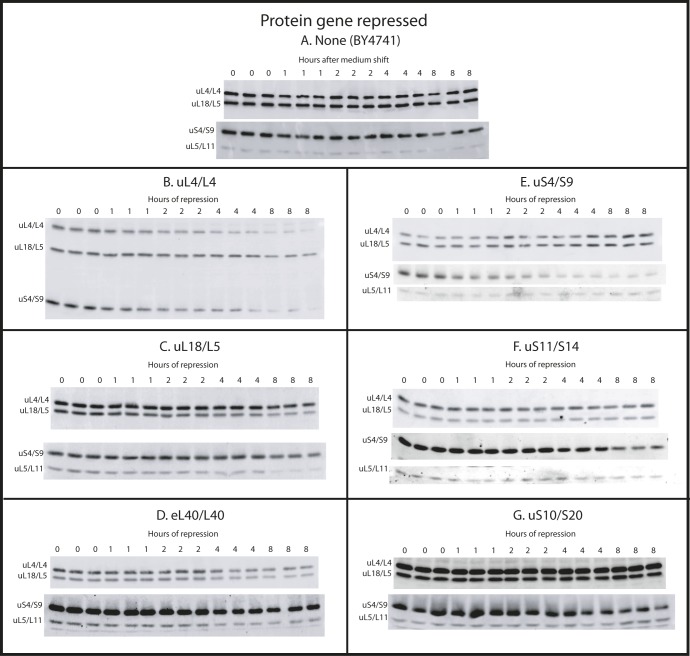
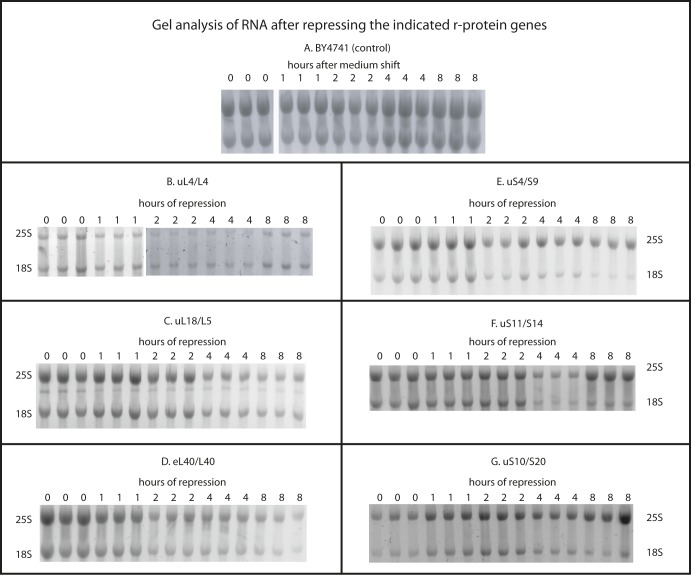
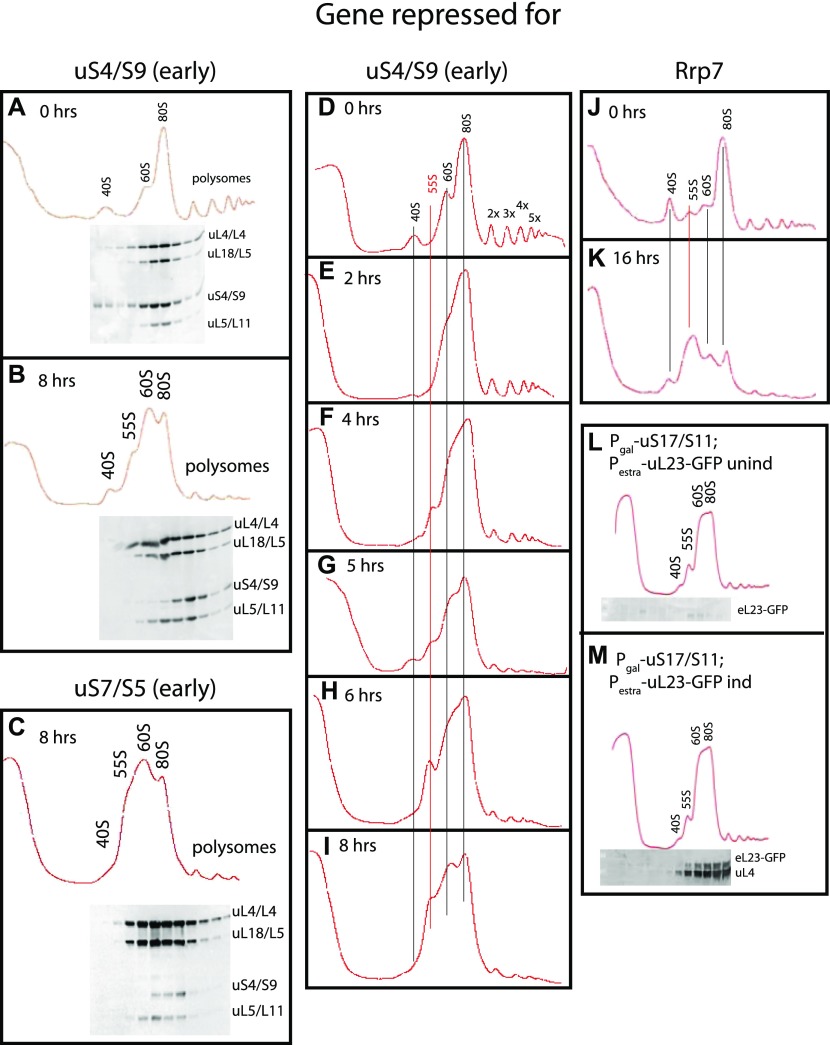
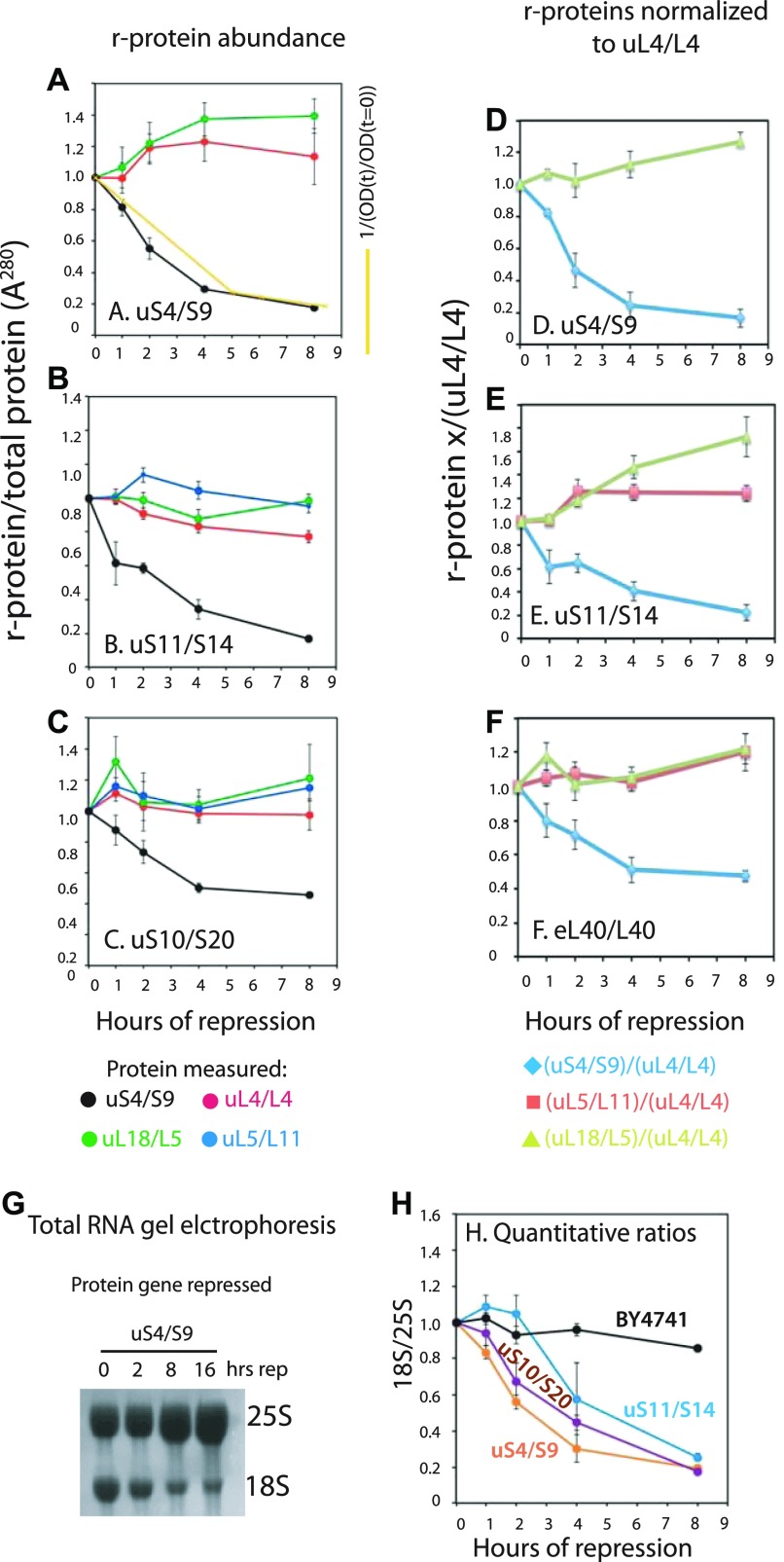
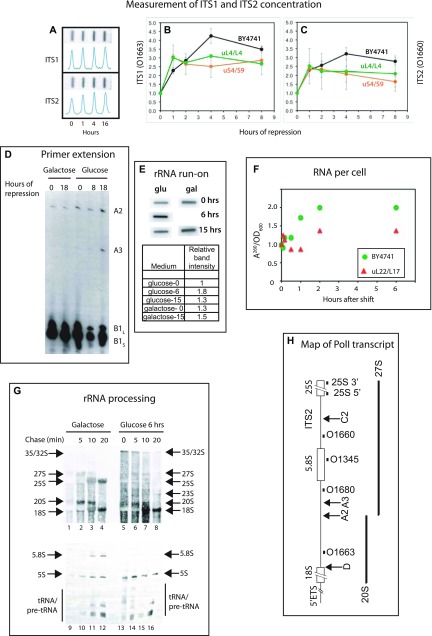
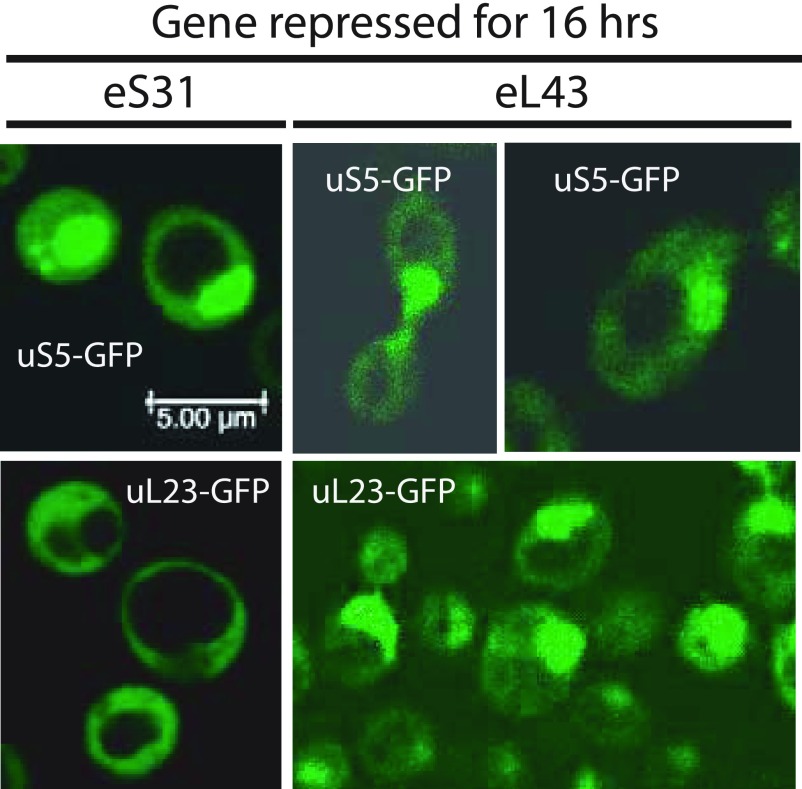
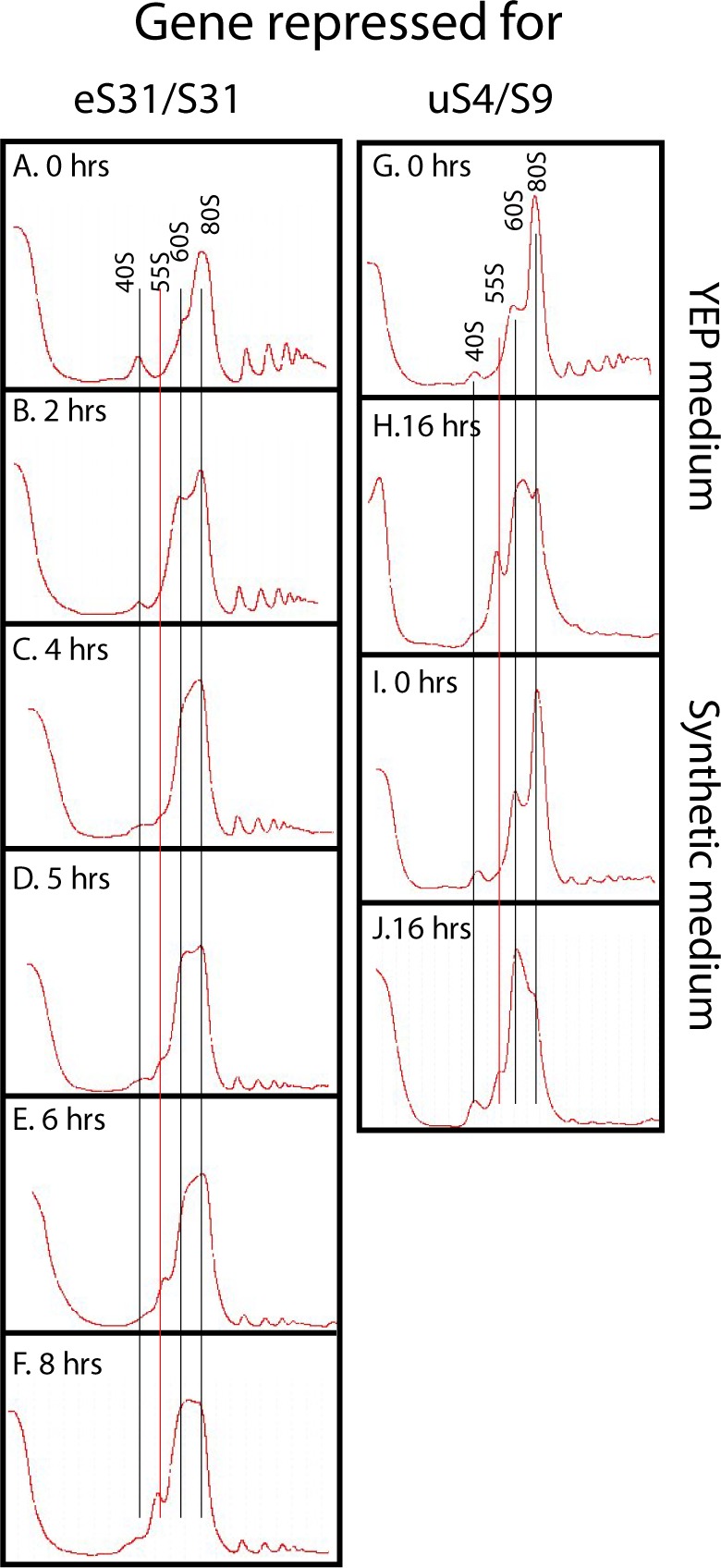
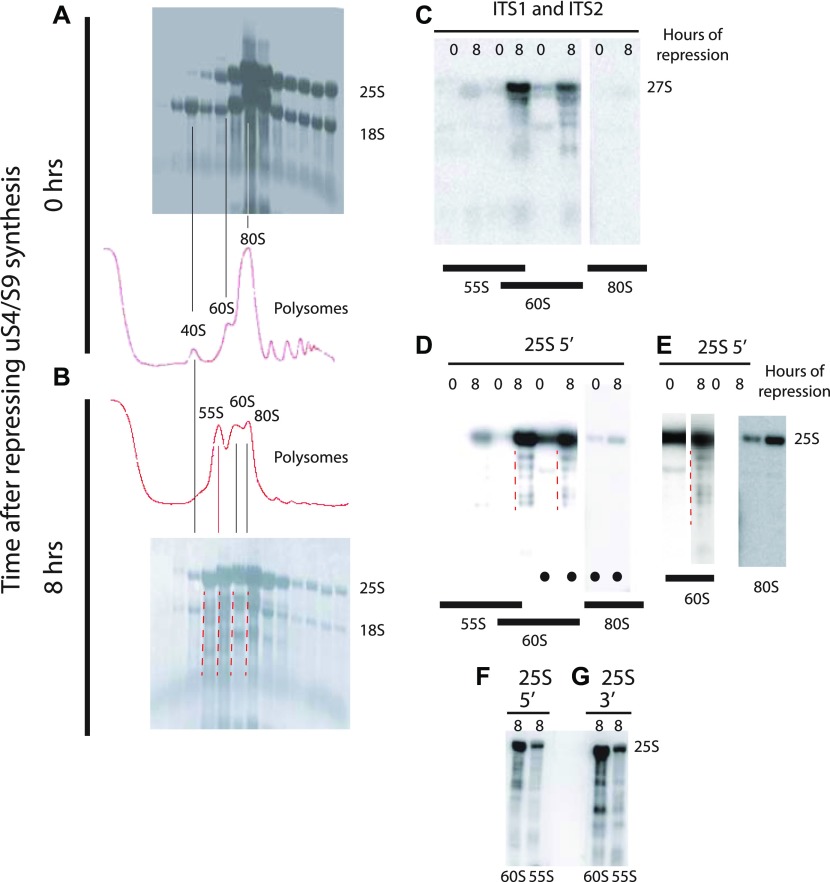
The 1:1 balance between the numbers of large and small ribosomal subunits can be disturbed by mutations that inhibit the assembly of only one of the subunits. Here, we have investigated if the cell can counteract an imbalance of the number of the two subunits. We show that abrogating 60S assembly blocks 40S subunit accumulation. In contrast, cessation of the 40S pathways does not prevent 60S accumulation, but does, however, lead to fragmentation of the 25S rRNA in 60S subunits and formation of a 55S ribosomal particle derived from the 60S. We also present evidence suggesting that these events occur post assembly and discuss the possibility that the turnover of subunits is due to vulnerability of free subunits not paired with the other subunit to form 80S ribosomes.
大、小核糖体亚基数量之间 1:1 的平衡可以被仅抑制一个亚基组装的突变所打破。在这里,我们研究了细胞是否可以抵消两个亚基数量的不平衡。我们表明,阻断 60S 组装会阻止 40S 亚基的积累。相比之下,停止 40S 途径不会阻止 60S 的积累,但确实导致 60S 亚基中 25S rRNA 的片段化,并形成源自 60S 的 55S 核糖体颗粒。我们还提供了表明这些事件发生在组装后发生的证据,并讨论了亚基周转率可能是由于与另一个亚基不成对的游离亚基易受影响,无法形成 80S 核糖体的可能性。