Oregon State University, Department of Biochemistry and Biophysics, 2011 Agricultural and Life Sciences, Corvallis, OR 97331, USA.
Department of Biochemistry, Institute for Protein Design, University of Washington, Molecular Engineering and Sciences Building, 4000 15th Ave NE, Seattle, WA 98195, USA.
J Mol Biol. 2020 Jul 24;432(16):4690-4704. doi: 10.1016/j.jmb.2020.06.014. Epub 2020 Jun 19.
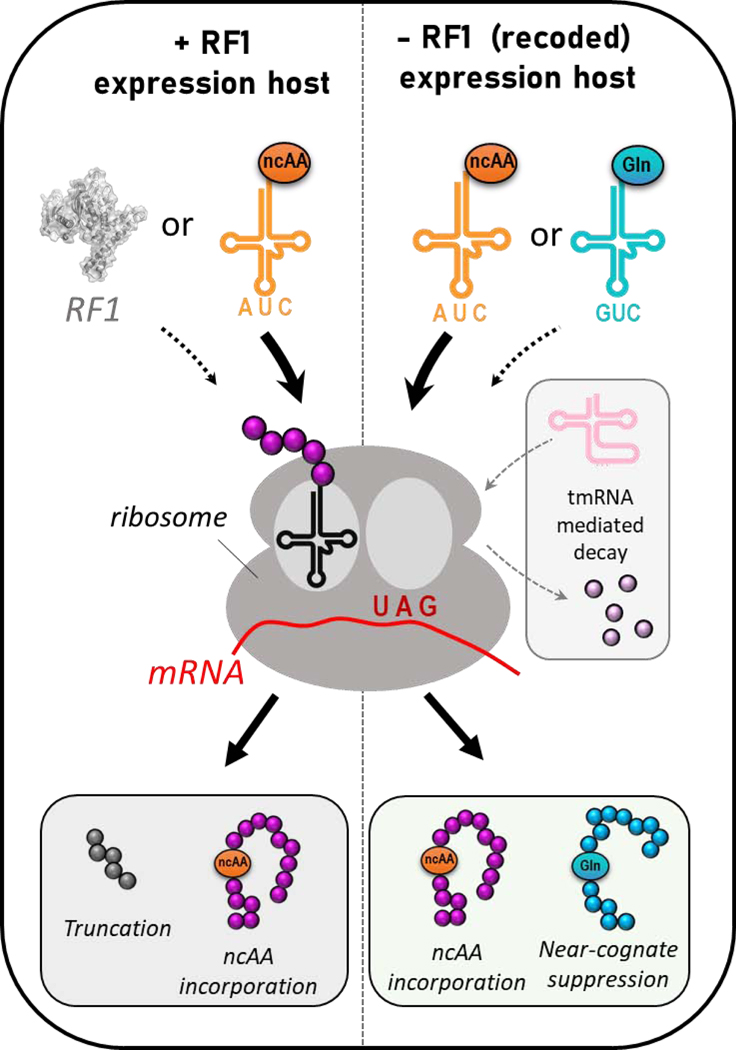
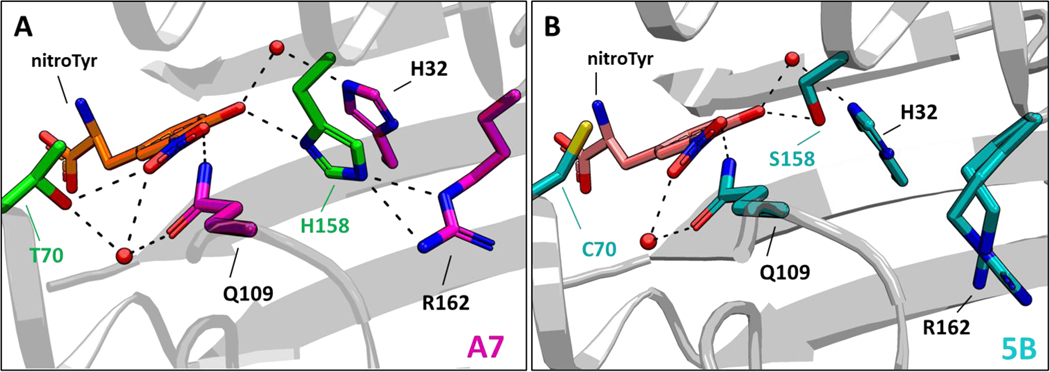
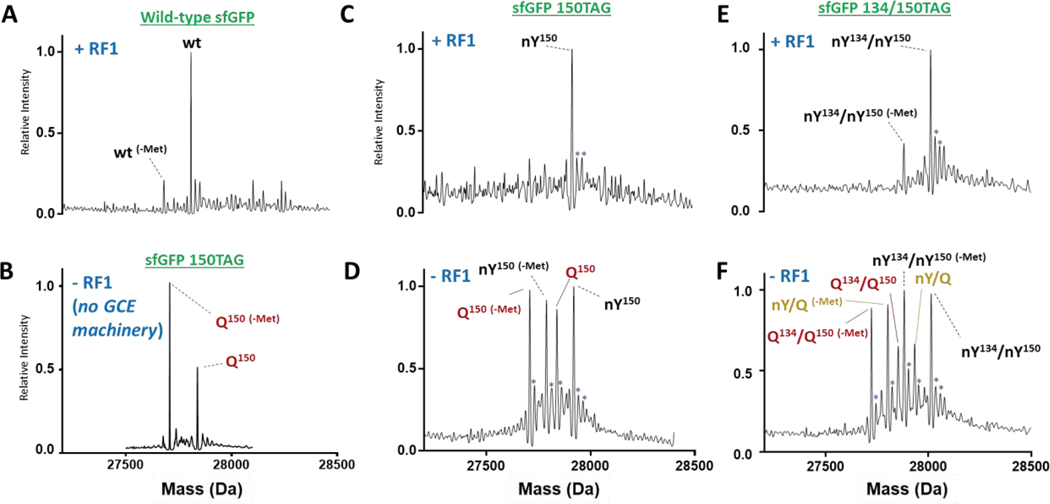
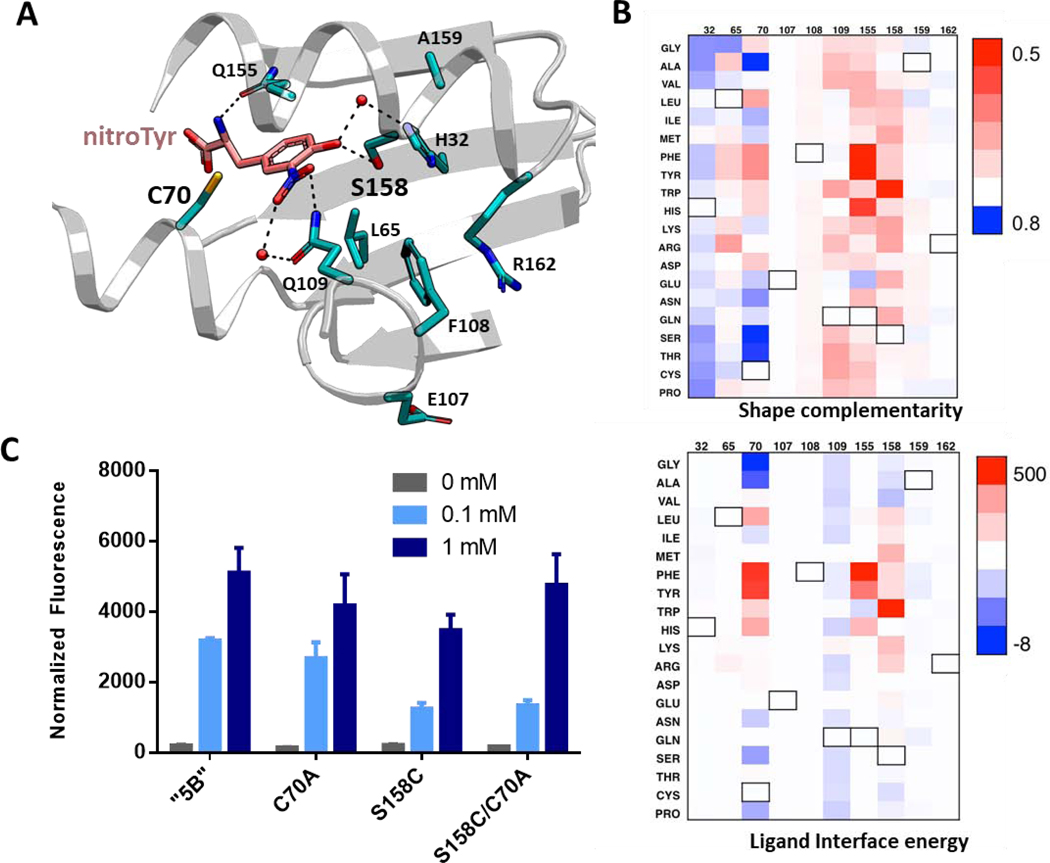
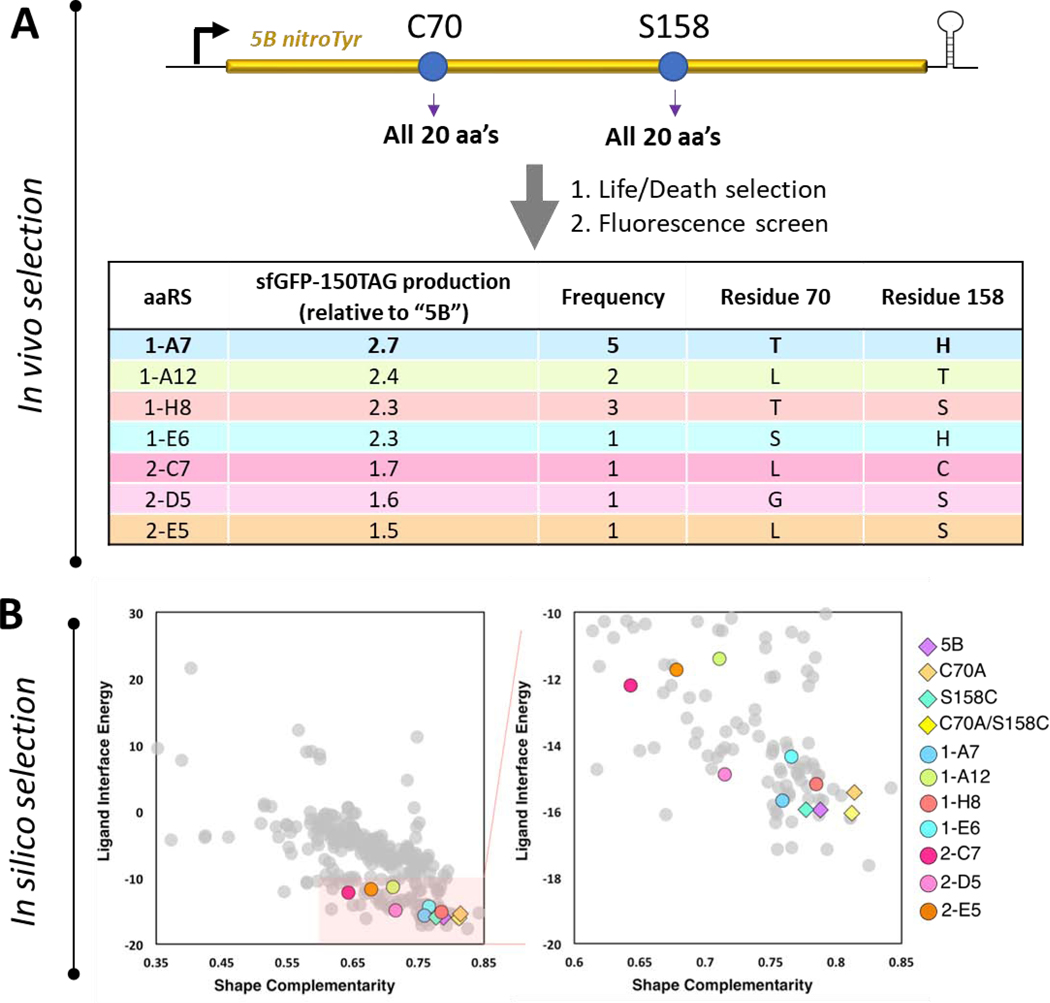
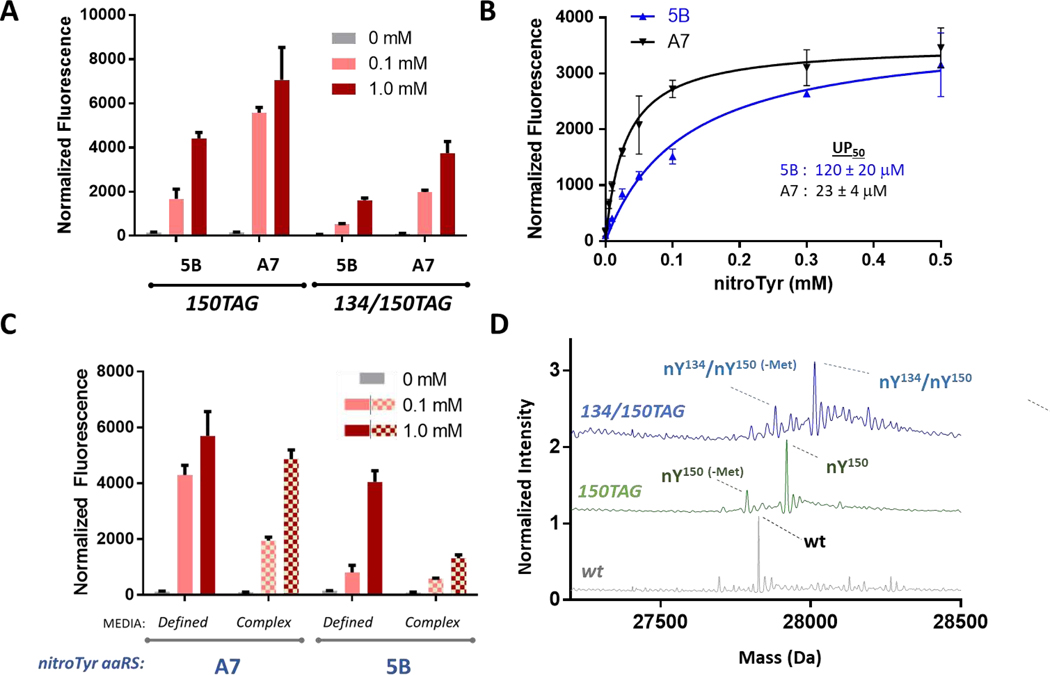
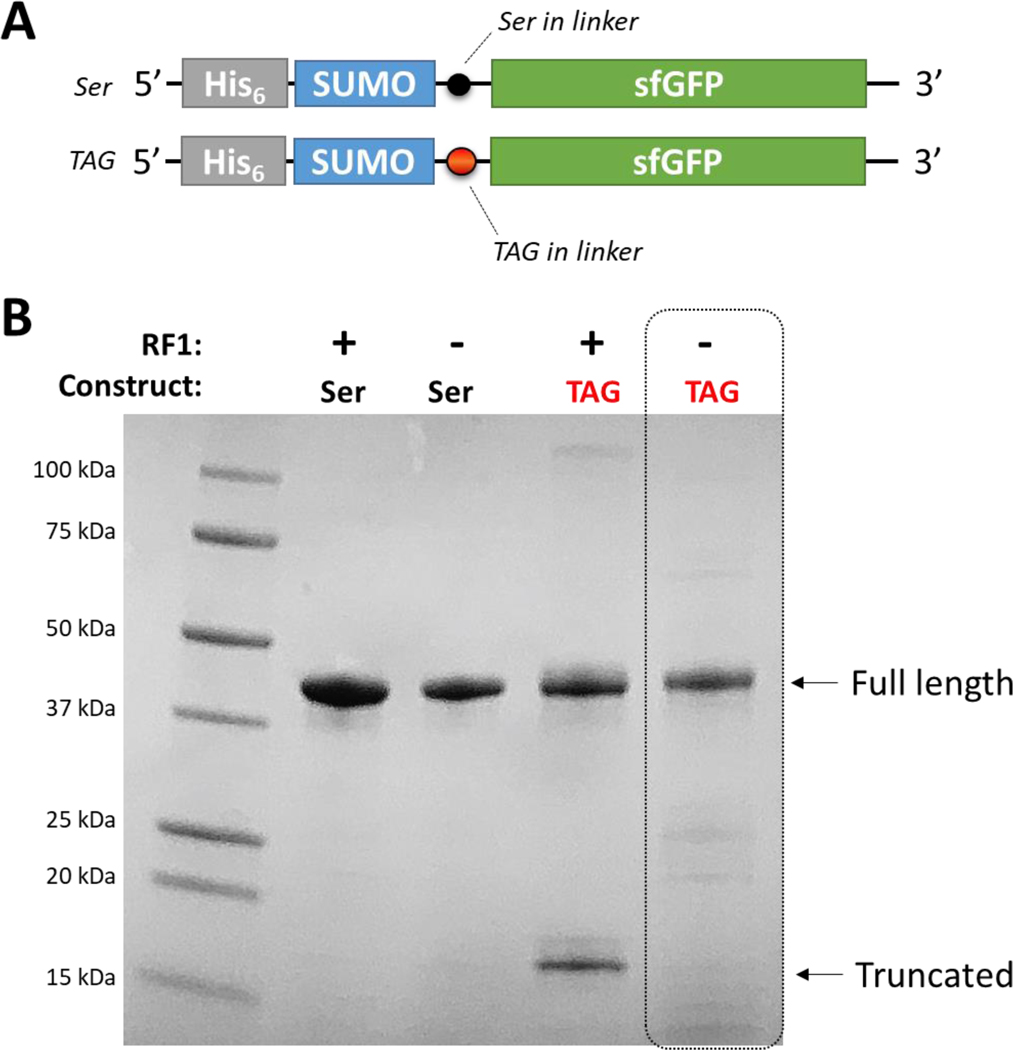
Genetic code expansion (GCE) technologies incorporate non-canonical amino acids (ncAAs) into proteins at amber stop codons. To avoid unwanted truncated protein and improve ncAA-protein yields, genomically recoded strains of Escherichia coli lacking Release Factor 1 (RF1) are becoming increasingly popular expression hosts for GCE applications. In the absence of RF1, however, endogenous near-cognate amber suppressing tRNAs can lead to contaminating protein forms with natural amino acids in place of the ncAA. Here, we show that a second-generation amino-acyl tRNA synthetase (aaRS)/tRNA pair for site-specific incorporation of 3-nitro-tyrosine could not outcompete near-cognate suppression in an RF1-deficient expression host and therefore could not produce homogenously nitrated protein. To resolve this, we used Rosetta to target positions in the nitroTyr aaRS active site for improved substrate binding, and then constructed of a small library of variants to subject to standard selection protocols. The top selected variant had an ~2-fold greater efficiency, and remarkably, this relatively small improvement enabled homogeneous incorporation of nitroTyr in an RF1-deficient expression host and thus eliminates truncation issues associated with typical RF1-containing expression hosts. Structural and biochemical data suggest the aaRS efficiency improvement is based on higher affinity substrate binding. Taken together, the modest improvement in aaRS efficiency provides a large practical impact and expands our ability to study the role protein nitration plays in disease development through producing homogenous, truncation-free nitroTyr-containing protein. This work establishes Rosetta-guided design and incremental aaRS improvement as a viable and accessible path to improve GCE systems challenged by truncation and/or near-cognate suppression issues.
遗传密码扩展(GCE)技术将非规范氨基酸(ncAA)掺入到琥珀终止密码子处的蛋白质中。为了避免产生不必要的截短蛋白并提高 ncAA-蛋白质的产量,缺乏释放因子 1(RF1)的基因重编码大肠杆菌菌株越来越成为 GCE 应用的流行表达宿主。然而,在没有 RF1 的情况下,内源性的近同功琥珀抑制 tRNA 可能导致含有天然氨基酸而不是 ncAA 的污染蛋白质形式。在这里,我们表明,用于定点掺入 3-硝基酪氨酸的第二代氨酰-tRNA 合成酶(aaRS)/tRNA 对不能在缺乏 RF1 的表达宿主中与近同功抑制竞争,因此不能产生均匀硝化的蛋白质。为了解决这个问题,我们使用 Rosetta 针对硝基 Tyr aaRS 活性位点中的位置进行了靶向,以改善底物结合,然后构建了一个小型变体文库,以进行标准选择方案。排名最高的变体具有约 2 倍的更高效率,值得注意的是,这种相对较小的改进使在缺乏 RF1 的表达宿主中均匀掺入硝基 Tyr 成为可能,从而消除了与典型含 RF1 的表达宿主相关的截断问题。结构和生化数据表明,aaRS 效率的提高基于更高的亲和力底物结合。总之,aaRS 效率的适度提高具有很大的实际影响,并扩大了我们通过产生均匀、无截短的硝基 Tyr 含有蛋白质来研究蛋白质硝化在疾病发展中所起作用的能力。这项工作确立了 Rosetta 引导的设计和 aaRS 的逐步改进作为一种可行且易于获得的途径,可以改善受到截断和/或近同功抑制问题挑战的 GCE 系统。