Department of Biochemistry and Biophysics, Stockholm University, SE 106 91 Stockholm, Sweden.
Center of Structural and Cell Biology in Medicine, Institute of Biochemistry, University of Lübeck, D-23538, Lübeck, Germany.
Chem Rev. 2021 May 12;121(9):5289-5335. doi: 10.1021/acs.chemrev.0c00983. Epub 2021 Apr 22.
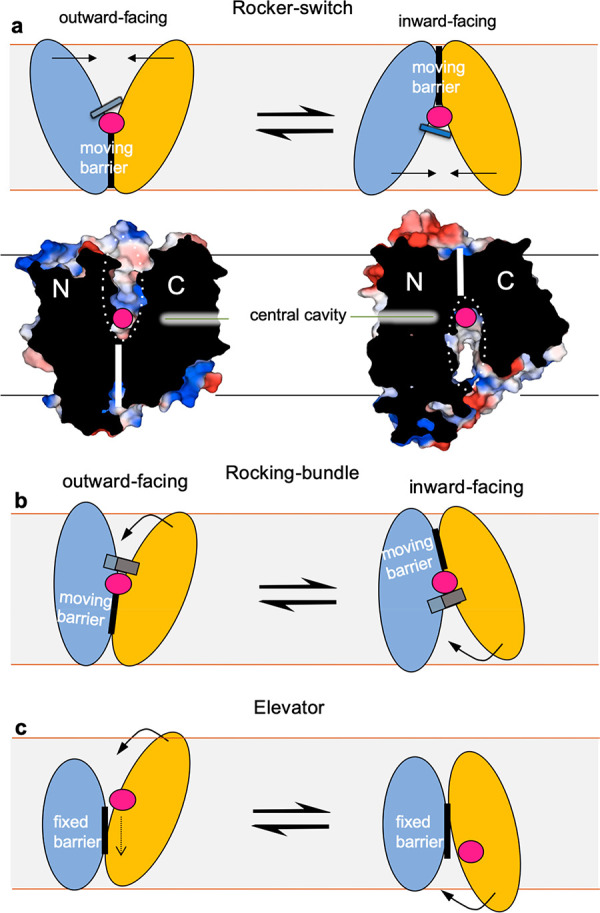
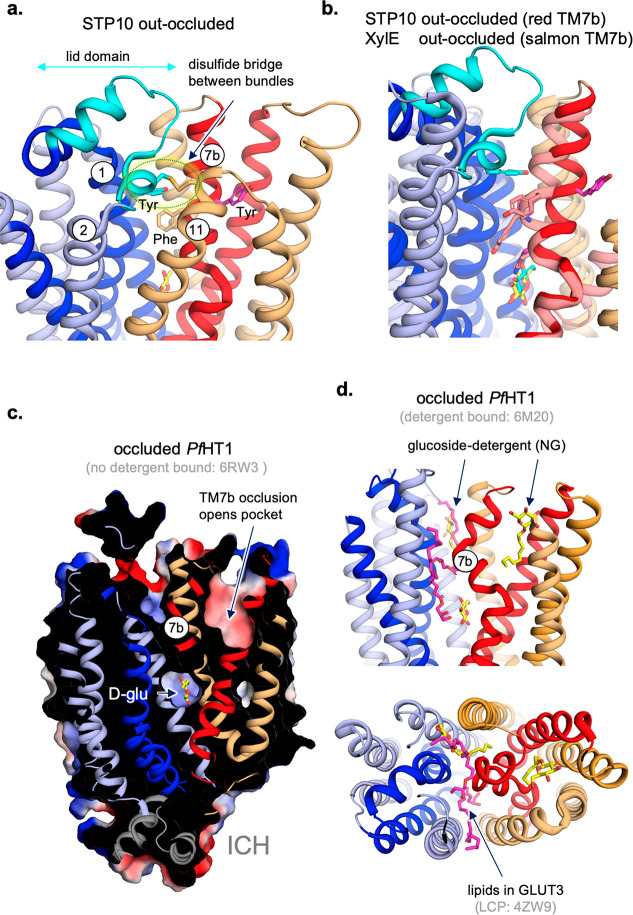
The major facilitator superfamily (MFS) is the largest known superfamily of secondary active transporters. MFS transporters are responsible for transporting a broad spectrum of substrates, either down their concentration gradient or uphill using the energy stored in the electrochemical gradients. Over the last 10 years, more than a hundred different MFS transporter structures covering close to 40 members have provided an atomic framework for piecing together the molecular basis of their transport cycles. Here, we summarize the remarkable promiscuity of MFS members in terms of substrate recognition and proton coupling as well as the intricate gating mechanisms undergone in achieving substrate translocation. We outline studies that show how residues far from the substrate binding site can be just as important for fine-tuning substrate recognition and specificity as those residues directly coordinating the substrate, and how a number of MFS transporters have evolved to form unique complexes with chaperone and signaling functions. Through a deeper mechanistic description of glucose (GLUT) transporters and multidrug resistance (MDR) antiporters, we outline novel refinements to the rocker-switch alternating-access model, such as a latch mechanism for proton-coupled monosaccharide transport. We emphasize that a full understanding of transport requires an elucidation of MFS transporter dynamics, energy landscapes, and the determination of how rate transitions are modulated by lipids.
主要促进因子超家族(MFS)是已知最大的次级主动转运蛋白超家族。MFS 转运蛋白负责运输广泛的底物,无论是沿着浓度梯度向下运输,还是利用电化学梯度中储存的能量向上运输。在过去的 10 年中,超过一百种不同的 MFS 转运蛋白结构涵盖了近 40 个成员,为拼凑它们的运输循环的分子基础提供了一个原子框架。在这里,我们总结了 MFS 成员在底物识别和质子偶联方面的显著混杂性,以及在实现底物转运过程中经历的复杂门控机制。我们概述了一些研究,这些研究表明远离底物结合位点的残基如何与直接协调底物的残基一样重要,以精细调整底物识别和特异性,以及许多 MFS 转运蛋白如何进化形成与伴侣蛋白和信号转导功能独特的复合物。通过对葡萄糖(GLUT)转运蛋白和多药耐药(MDR)外排泵的更深入的机制描述,我们概述了 rocker-switch 交替访问模型的新改进,例如用于质子偶联单糖转运的闩锁机制。我们强调,要全面了解运输,需要阐明 MFS 转运蛋白的动力学、能量景观,以及确定如何通过脂质调节速率跃迁。