Department of Biology, Massachusetts Institute of Technology, Cambridge, United States.
David H. Koch Institute for Integrative Cancer Research, Massachusetts Institute of Technology, Cambridge, United States.
Elife. 2022 Sep 13;11:e77058. doi: 10.7554/eLife.77058.
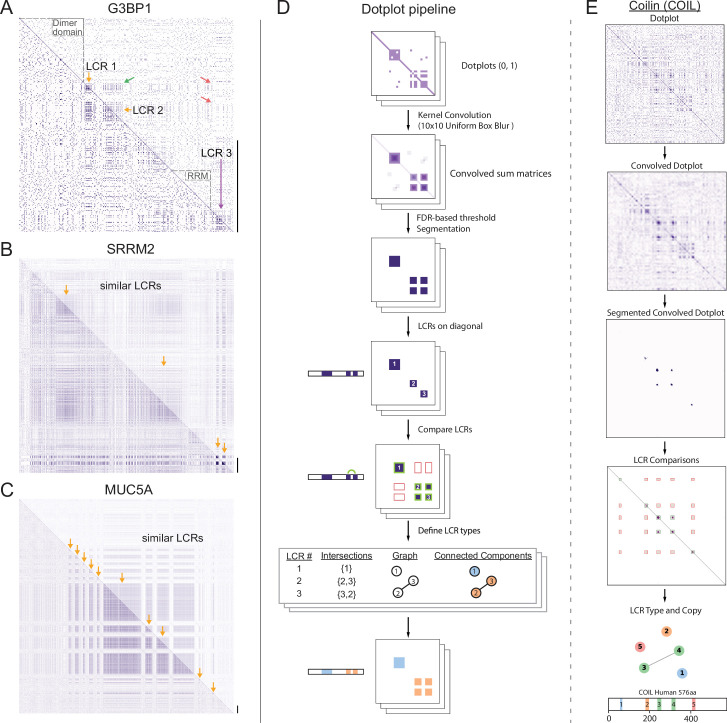
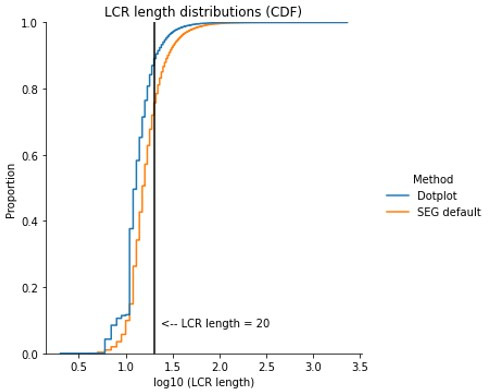
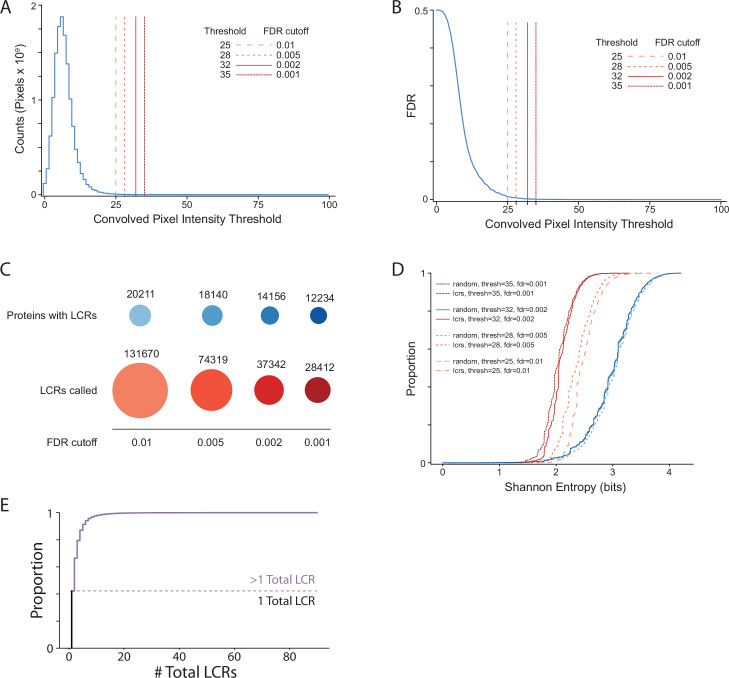
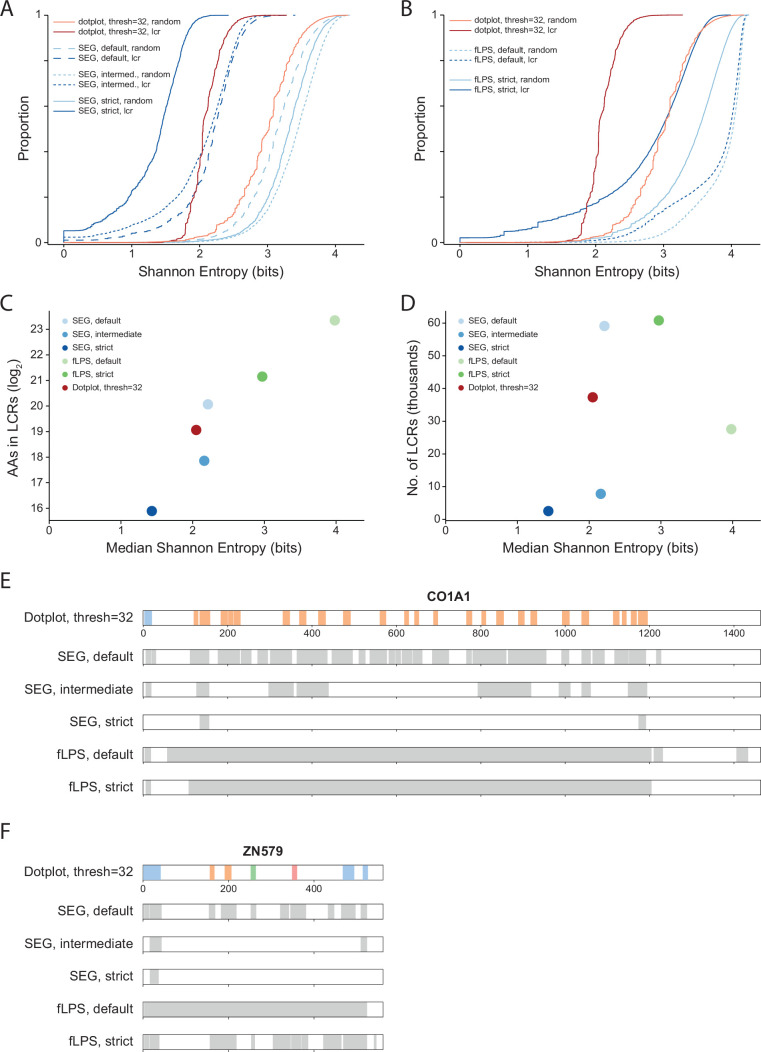
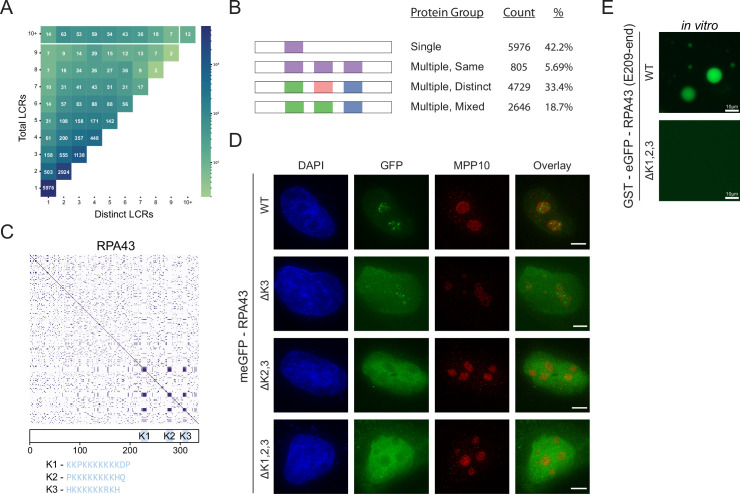
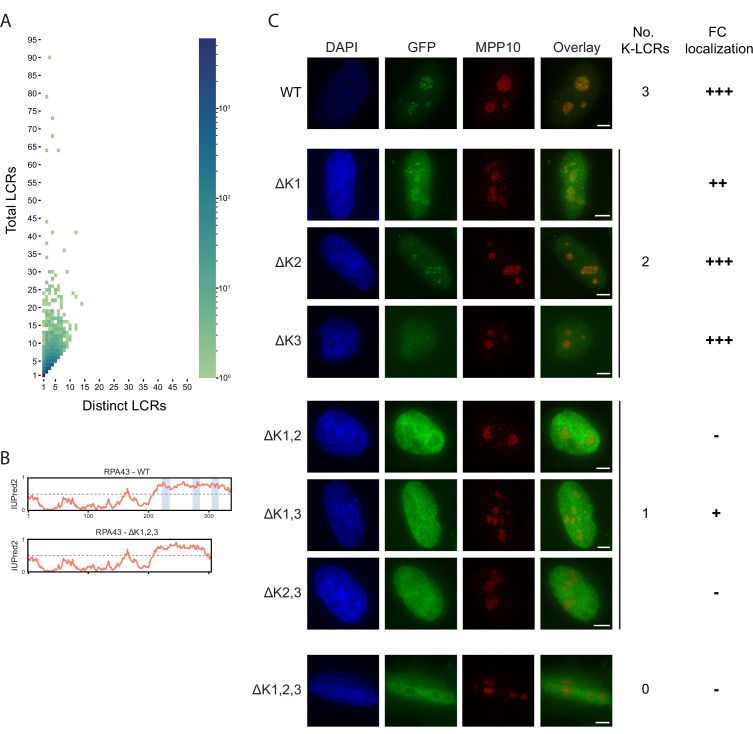
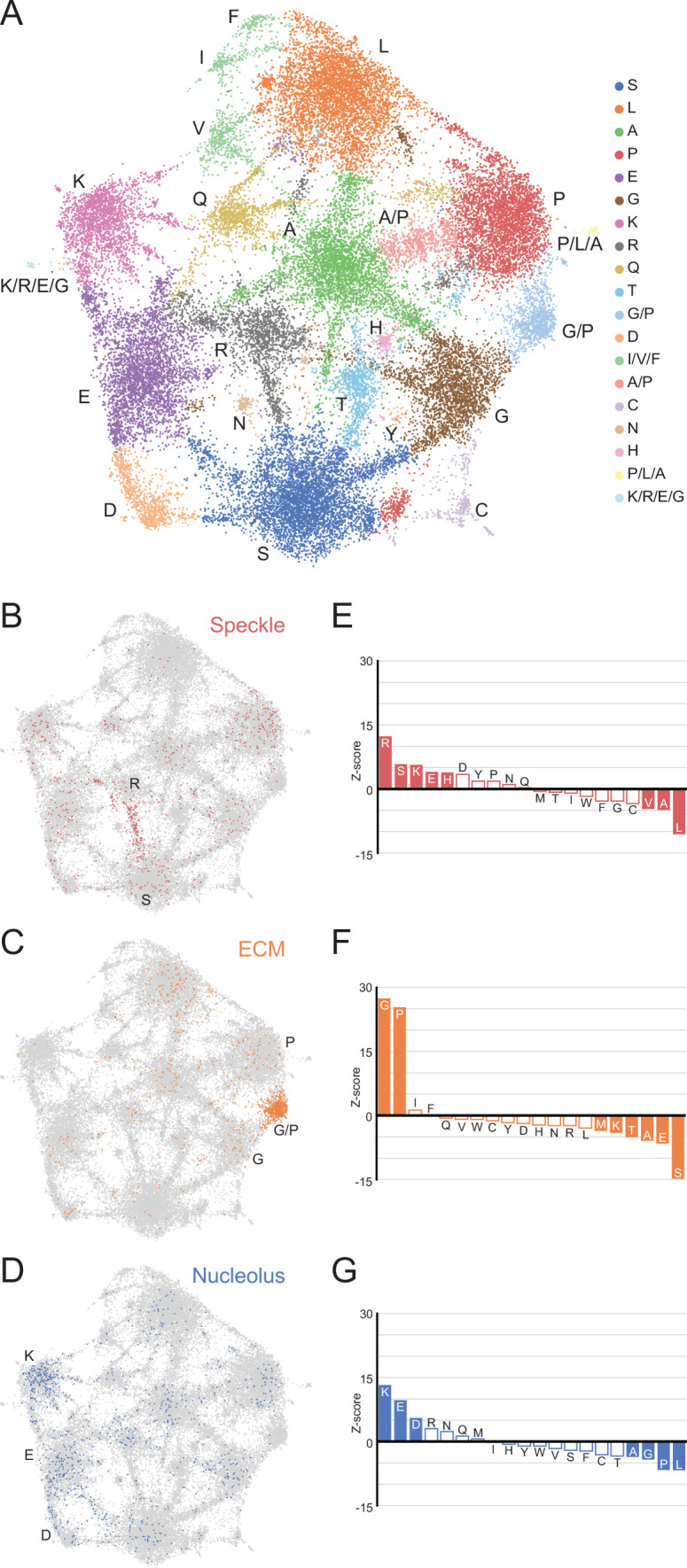
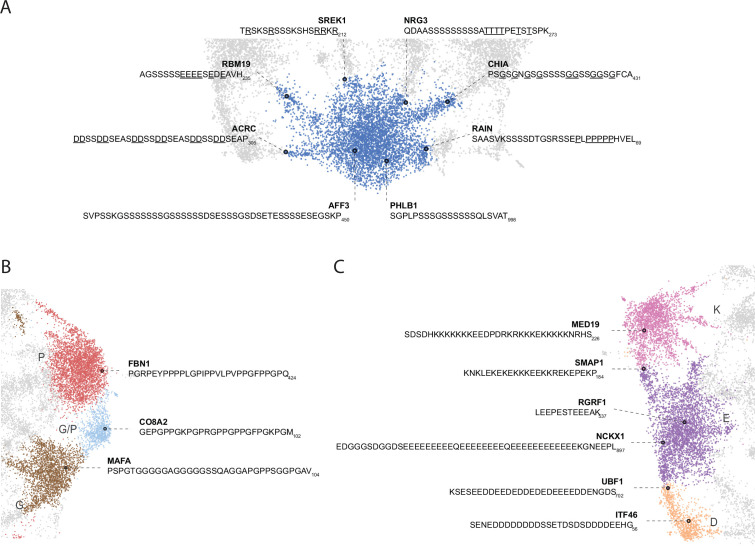
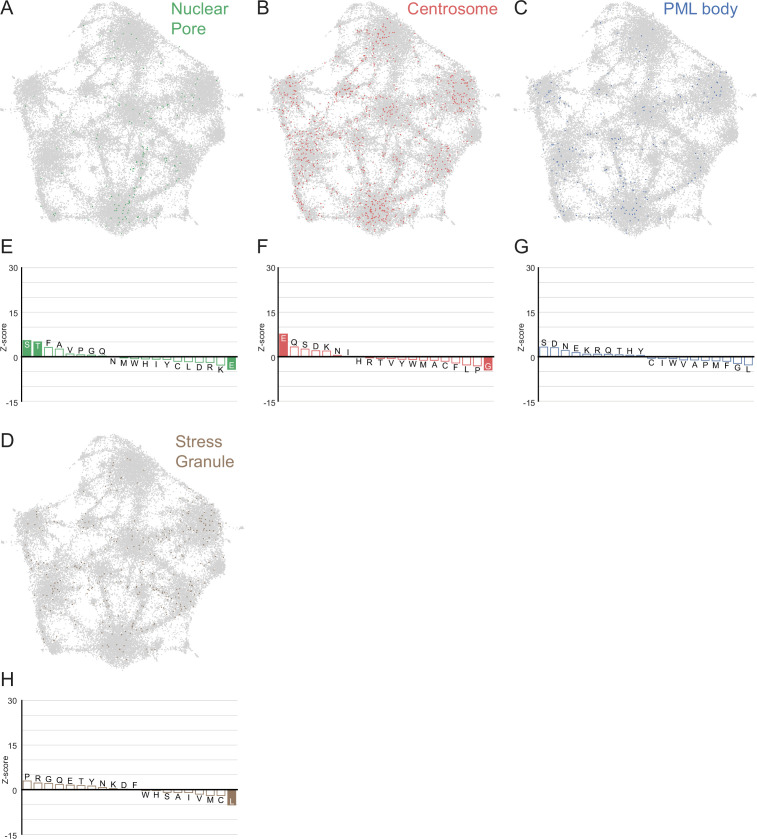
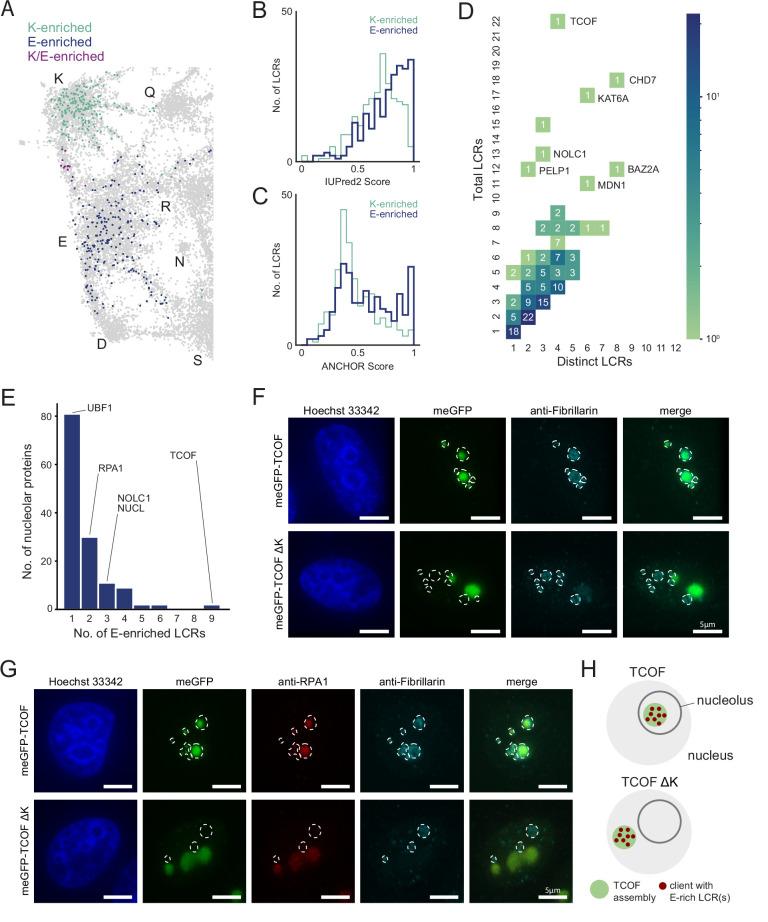
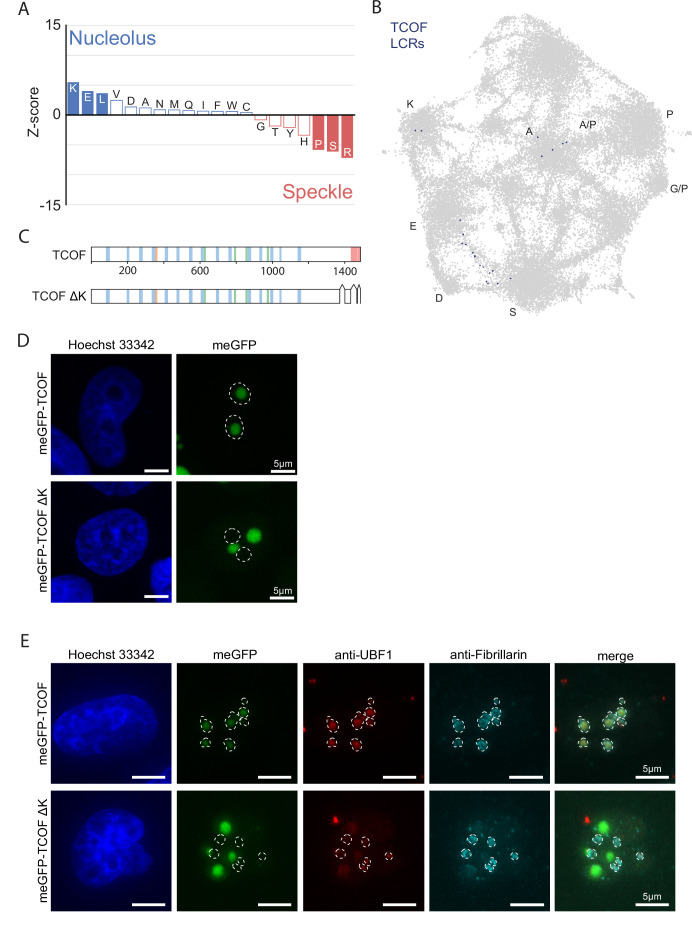
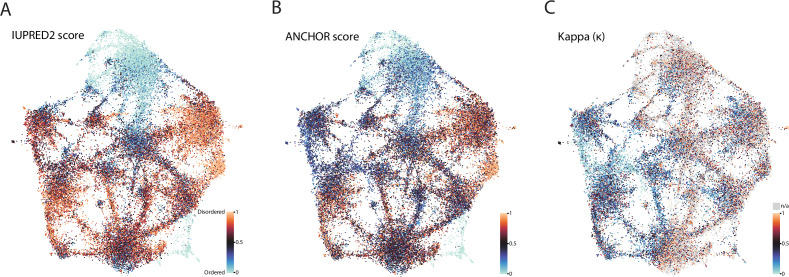
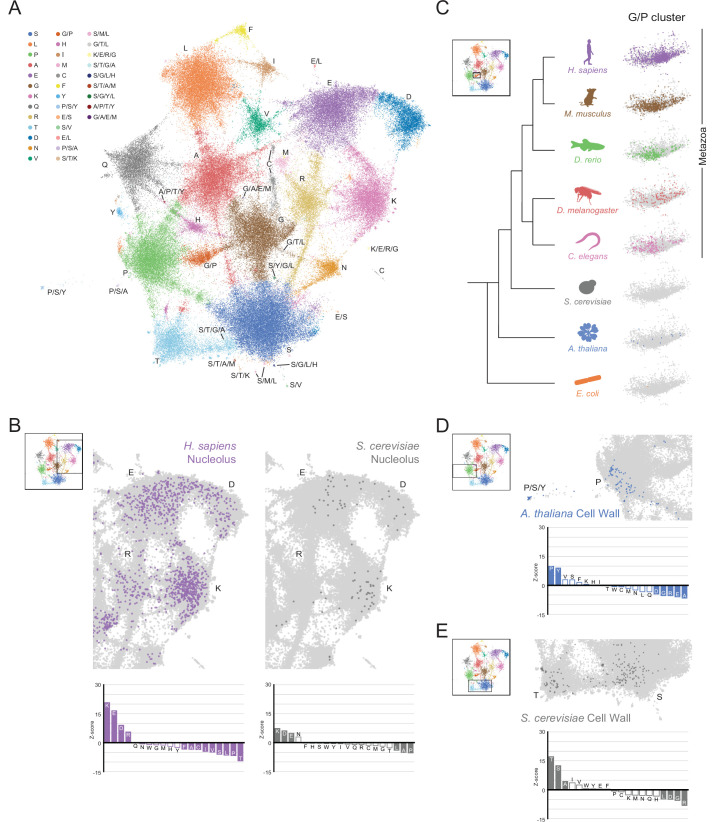
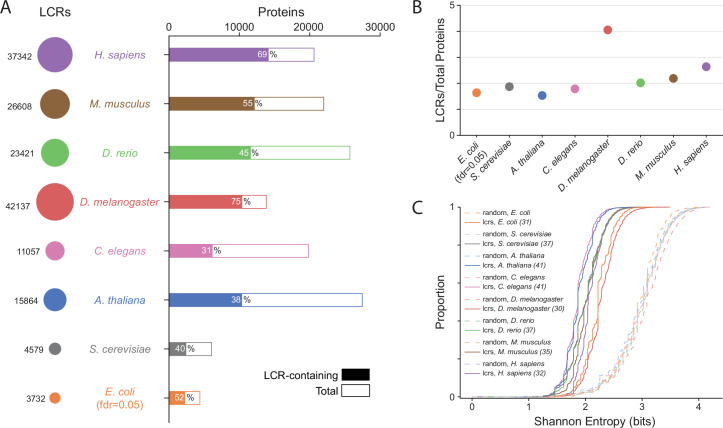
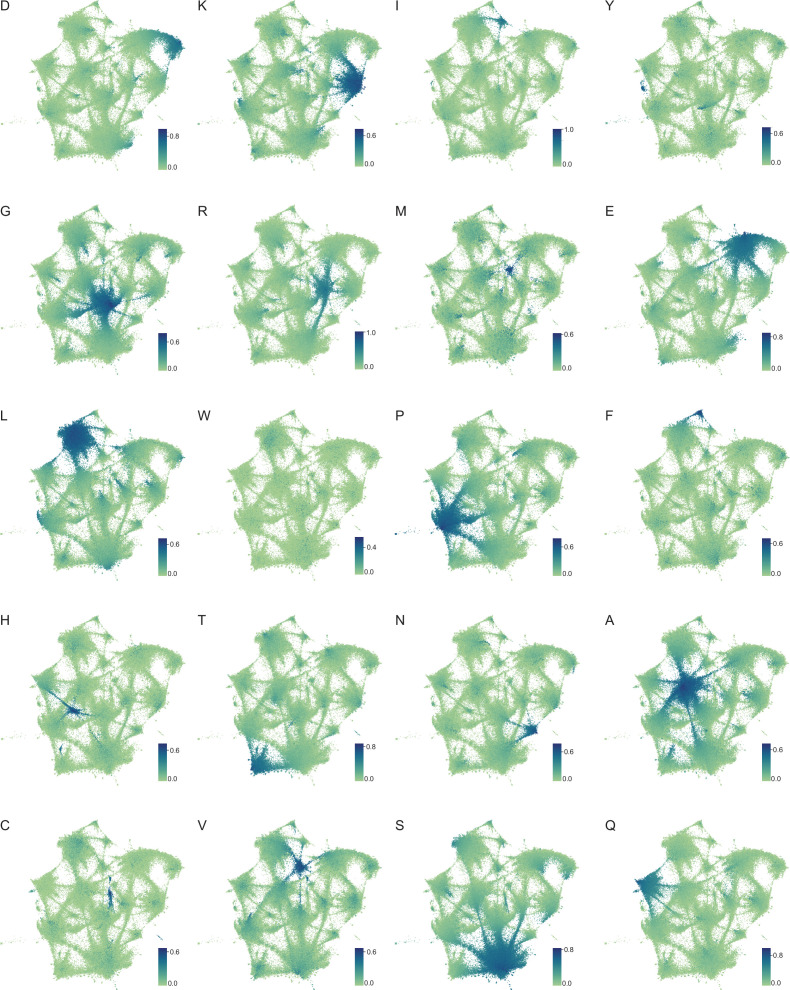
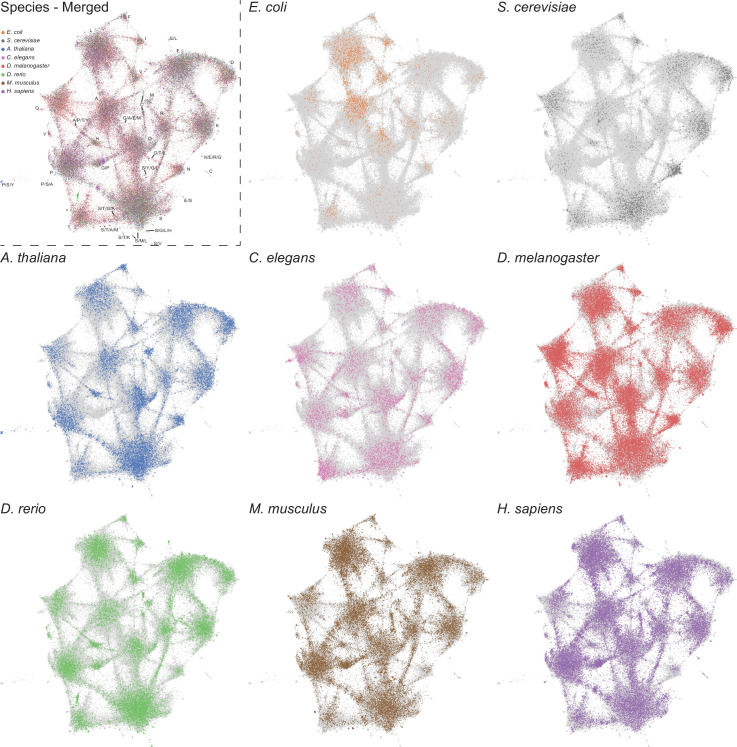
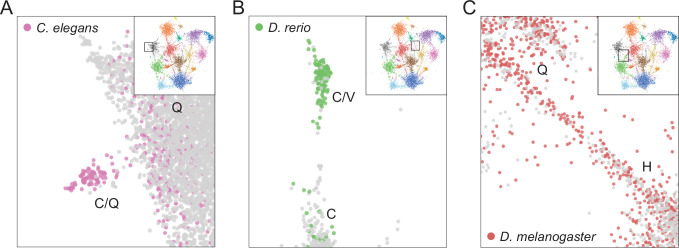
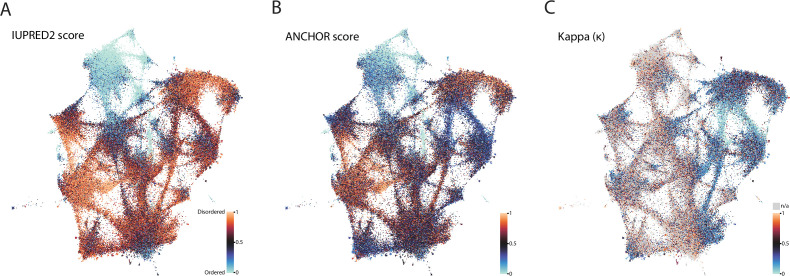
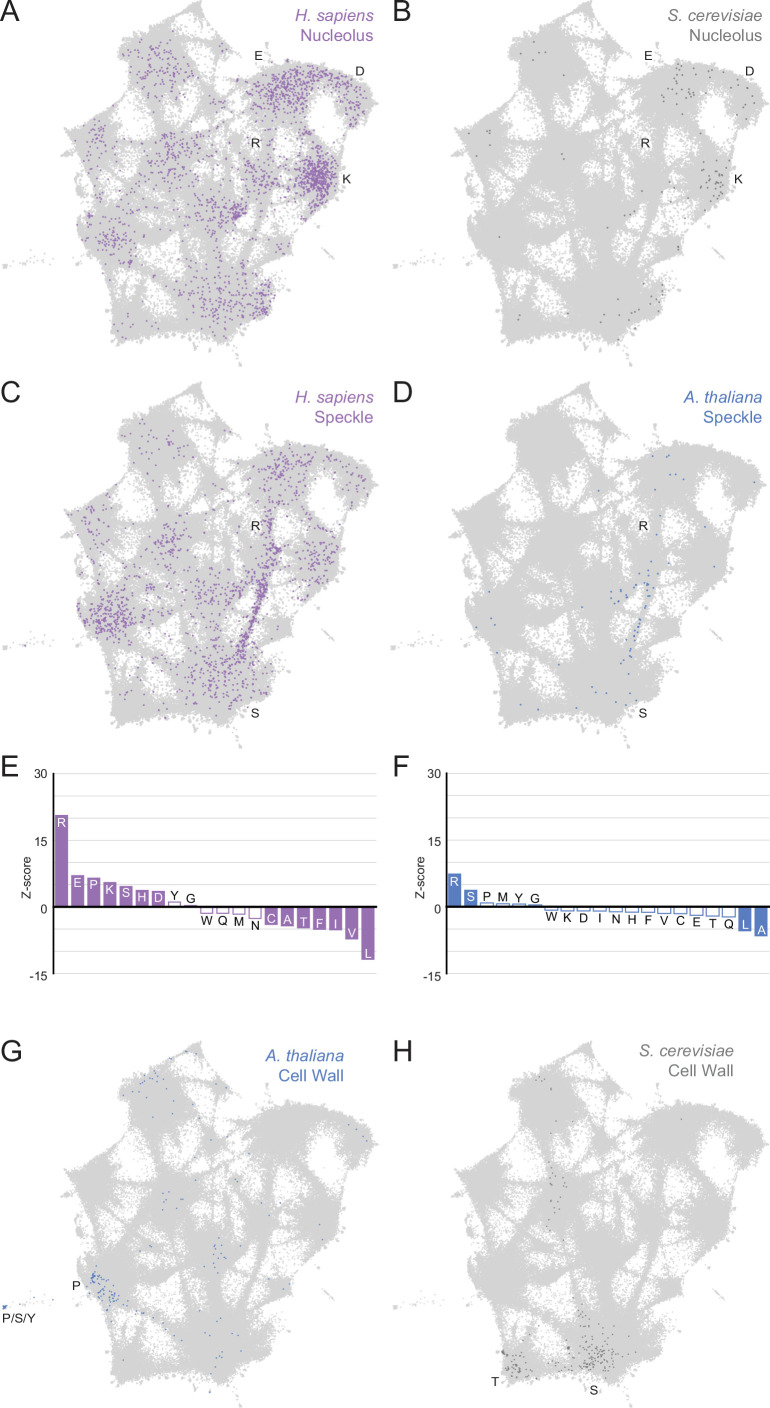
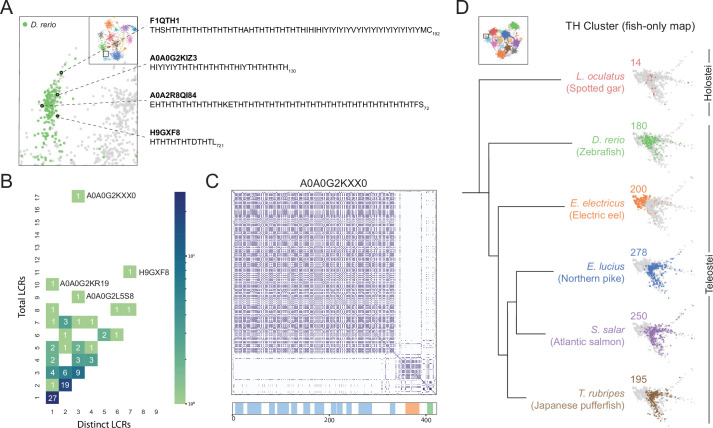
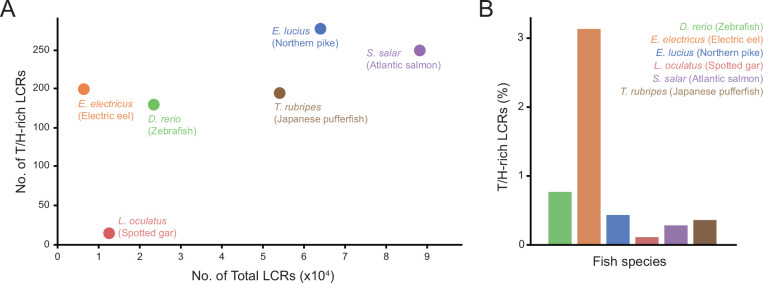
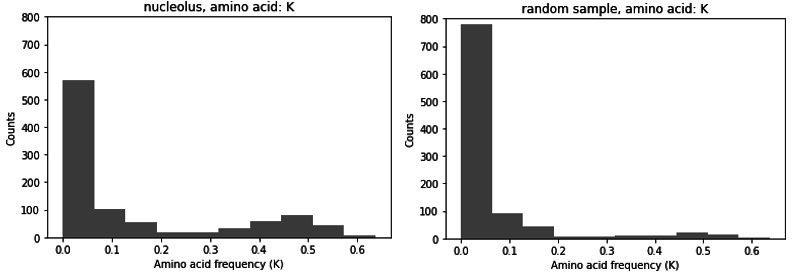
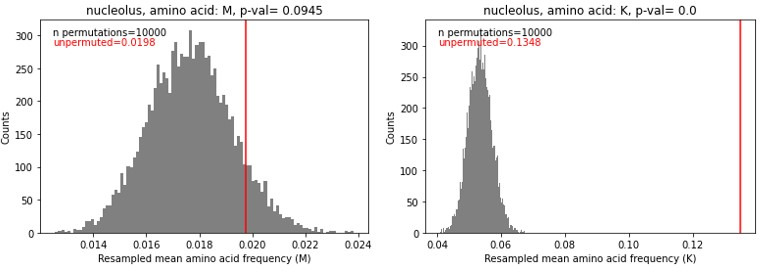
Low complexity regions (LCRs) play a role in a variety of important biological processes, yet we lack a unified view of their sequences, features, relationships, and functions. Here, we use dotplots and dimensionality reduction to systematically define LCR type/copy relationships and create a map of LCR sequence space capable of integrating LCR features and functions. By defining LCR relationships across the proteome, we provide insight into how LCR type and copy number contribute to higher order assemblies, such as the importance of K-rich LCR copy number for assembly of the nucleolar protein RPA43 in vivo and in vitro. With LCR maps, we reveal the underlying structure of LCR sequence space, and relate differential occupancy in this space to the conservation and emergence of higher order assemblies, including the metazoan extracellular matrix and plant cell wall. Together, LCR relationships and maps uncover and identify scaffold-client relationships among E-rich LCR-containing proteins in the nucleolus, and revealed previously undescribed regions of LCR sequence space with signatures of higher order assemblies, including a teleost-specific T/H-rich sequence space. Thus, this unified view of LCRs enables discovery of how LCRs encode higher order assemblies of organisms.
低复杂度区域(LCRs)在多种重要的生物学过程中发挥作用,但我们缺乏对它们的序列、特征、关系和功能的统一认识。在这里,我们使用点图和降维来系统地定义 LCR 类型/拷贝关系,并创建一个能够整合 LCR 特征和功能的 LCR 序列空间图谱。通过在整个蛋白质组中定义 LCR 关系,我们深入了解了 LCR 类型和拷贝数如何为更高阶的组装做出贡献,例如富含 K 的 LCR 拷贝数对体内和体外核仁蛋白 RPA43 组装的重要性。通过 LCR 图谱,我们揭示了 LCR 序列空间的潜在结构,并将其与空间中差异占据的关系与更高阶组装的保守性和出现联系起来,包括后生动物细胞外基质和植物细胞壁。总之,LCR 关系和图谱揭示并鉴定了核仁中富含 E 的 LCR 蛋白之间的支架-客户关系,并揭示了以前未描述的具有更高阶组装特征的 LCR 序列空间区域,包括一个硬骨鱼特异性的 T/H-富含序列空间。因此,这种对 LCR 的统一观点使我们能够发现 LCR 如何编码生物体的更高阶组装。