Walker Emma J, Hamill Carlin J, Crean Rory, Connolly Michael S, Warrender Annmaree K, Kraakman Kirsty L, Prentice Erica J, Steyn-Ross Alistair, Steyn-Ross Moira, Pudney Christopher R, van der Kamp Marc W, Schipper Louis A, Mulholland Adrian J, Arcus Vickery L
Te Aka Ma̅tuatua School of Science, University of Waikato, Hamilton 3214, New Zealand.
Centre for Computational Chemistry, School of Chemistry, University of Bristol, Bristol BS8 1TS, U.K.
ACS Catal. 2024 Mar 8;14(7):4379-4394. doi: 10.1021/acscatal.3c05584. eCollection 2024 Apr 5.
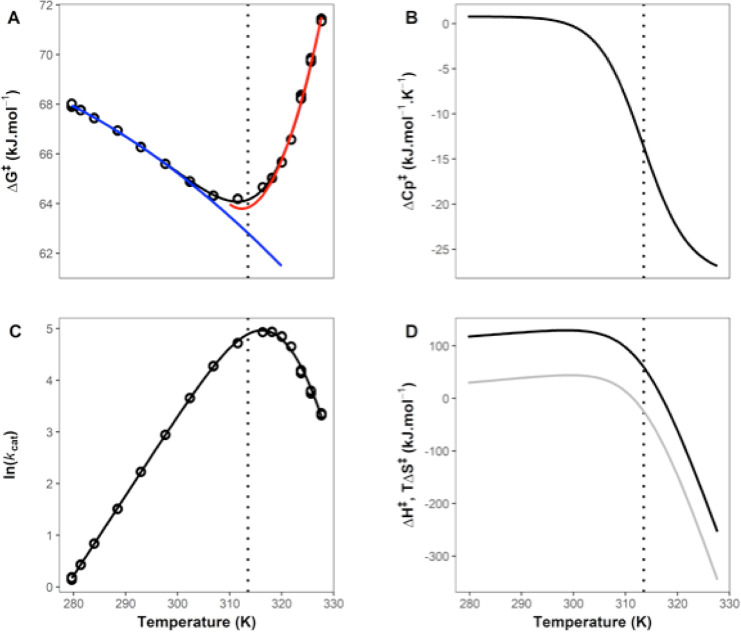
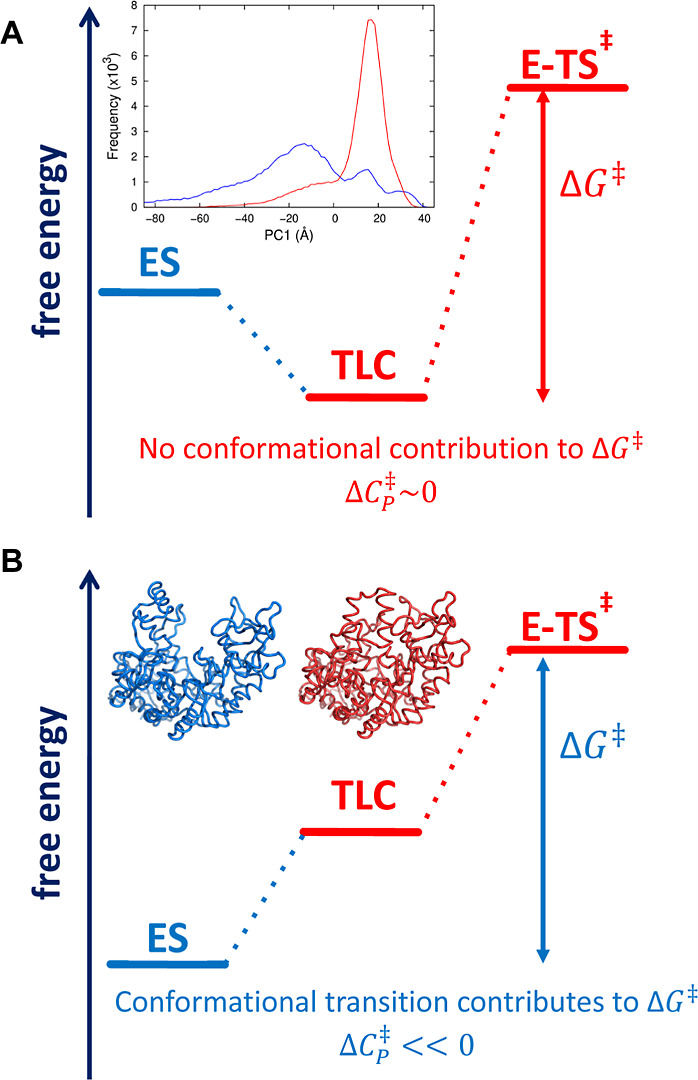
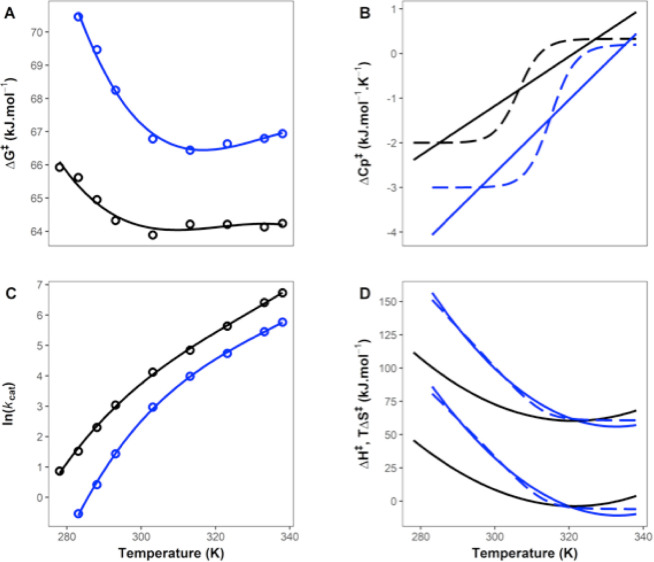
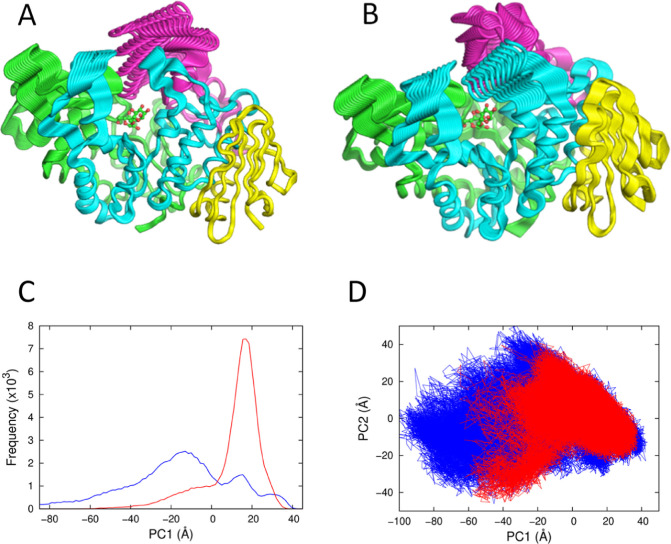
Many enzymes display non-Arrhenius behavior with curved Arrhenius plots in the absence of denaturation. There has been significant debate about the origin of this behavior and recently the role of the activation heat capacity (Δ) has been widely discussed. If enzyme-catalyzed reactions occur with appreciable negative values of Δ (arising from narrowing of the conformational space along the reaction coordinate), then curved Arrhenius plots are a consequence. To investigate these phenomena in detail, we have collected high precision temperature-rate data over a wide temperature interval for a model glycosidase enzyme MalL, and a series of mutants that change the temperature-dependence of the enzyme-catalyzed rate. We use these data to test a range of models including macromolecular rate theory (MMRT) and an equilibrium model. In addition, we have performed extensive molecular dynamics (MD) simulations to characterize the conformational landscape traversed by MalL in the enzyme-substrate complex and an enzyme-transition state complex. We have crystallized the enzyme in a transition state-like conformation in the absence of a ligand and determined an X-ray crystal structure at very high resolution (1.10 Å). We show (using simulation) that this enzyme-transition state conformation has a more restricted conformational landscape than the wildtype enzyme. We coin the term "transition state-like conformation (TLC)" to apply to this state of the enzyme. Together, these results imply a cooperative conformational transition between an enzyme-substrate conformation (ES) and a transition-state-like conformation (TLC) that precedes the chemical step. We present a two-state model as an extension of MMRT (MMRT-2S) that describes the data along with a convenient approximation with linear temperature dependence of the activation heat capacity (MMRT-1L) that can be used where fewer data points are available. Our model rationalizes disparate behavior seen for MalL and previous results for a thermophilic alcohol dehydrogenase and is consistent with a raft of data for other enzymes. Our model can be used to characterize the conformational changes required for enzyme catalysis and provides insights into the role of cooperative conformational changes in transition state stabilization that are accompanied by changes in heat capacity for the system along the reaction coordinate. TLCs are likely to be of wide importance in understanding the temperature dependence of enzyme activity and other aspects of enzyme catalysis.
许多酶在未发生变性时,其阿累尼乌斯图呈曲线,表现出非阿累尼乌斯行为。关于这种行为的起源存在大量争论,最近活化热容(Δ)的作用受到了广泛讨论。如果酶催化反应发生时Δ具有显著的负值(这是由于沿反应坐标的构象空间变窄所致),那么阿累尼乌斯图呈曲线就是其结果。为了详细研究这些现象,我们在很宽的温度区间内收集了模型糖苷酶MalL以及一系列改变酶催化速率温度依赖性的突变体的高精度温度 - 速率数据。我们用这些数据来测试一系列模型,包括大分子速率理论(MMRT)和一个平衡模型。此外,我们进行了广泛的分子动力学(MD)模拟,以表征MalL在酶 - 底物复合物和酶 - 过渡态复合物中所经历的构象景观。我们在没有配体的情况下使该酶结晶为类似过渡态的构象,并以非常高的分辨率(1.10 Å)确定了X射线晶体结构。我们通过模拟表明,这种酶 - 过渡态构象比野生型酶具有更受限的构象景观。我们创造了“类似过渡态构象(TLC)”一词来描述酶的这种状态。这些结果共同表明,在化学步骤之前,酶 - 底物构象(ES)和类似过渡态的构象(TLC)之间存在协同构象转变。我们提出了一个双态模型作为MMRT的扩展(MMRT - 2S),它可以描述这些数据,同时还提出了一种方便的近似方法,即活化热容具有线性温度依赖性(MMRT - 1L),可用于数据点较少的情况。我们的模型解释了MalL以及先前嗜热醇脱氢酶所呈现的不同行为,并与其他酶的大量数据一致。我们的模型可用于表征酶催化所需的构象变化,并深入了解协同构象变化在过渡态稳定化中的作用,这种稳定化伴随着系统沿反应坐标的热容变化。TLC在理解酶活性的温度依赖性以及酶催化的其他方面可能具有广泛的重要性。