TI Food & Nutrition, Nieuwe Kanaal 9A, 6709 PA, Wageningen, The Netherlands.
Microb Cell Fact. 2012 Sep 11;11:123. doi: 10.1186/1475-2859-11-123.
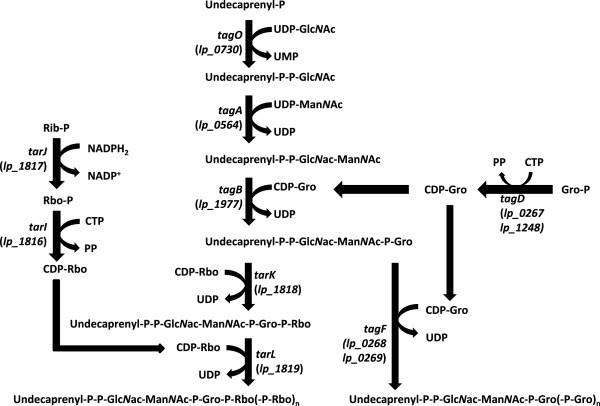
Specific strains of Lactobacillus plantarum are marketed as health-promoting probiotics. The role and interplay of cell-wall compounds like wall- and lipo-teichoic acids (WTA and LTA) in bacterial physiology and probiotic-host interactions remain obscure. L. plantarum WCFS1 harbors the genetic potential to switch WTA backbone alditol, providing an opportunity to study the impact of WTA backbone modifications in an isogenic background.
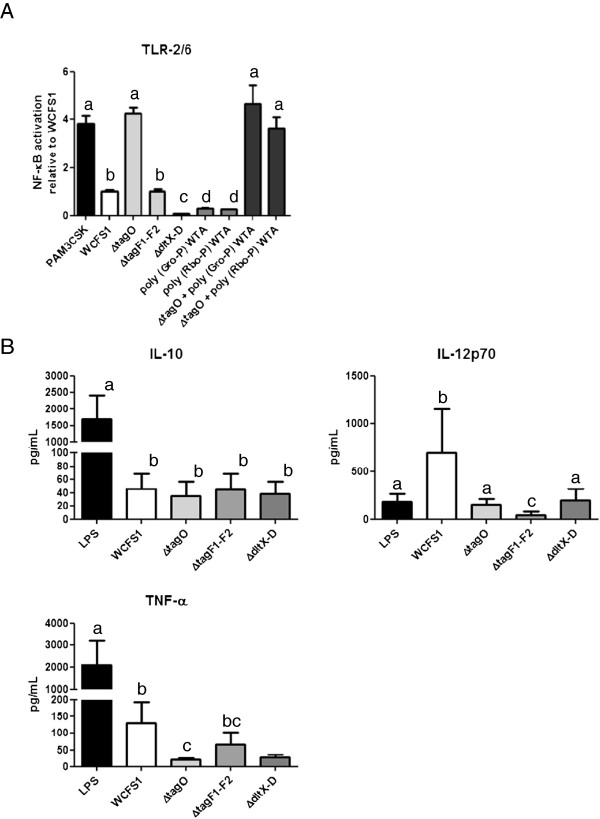
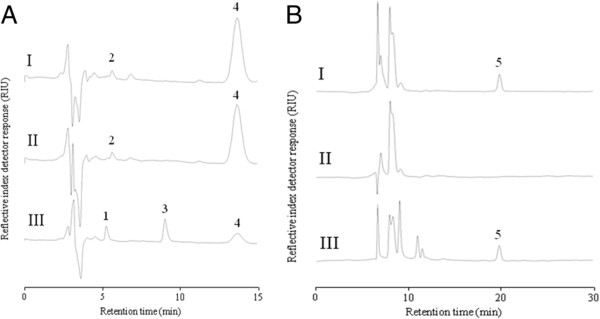
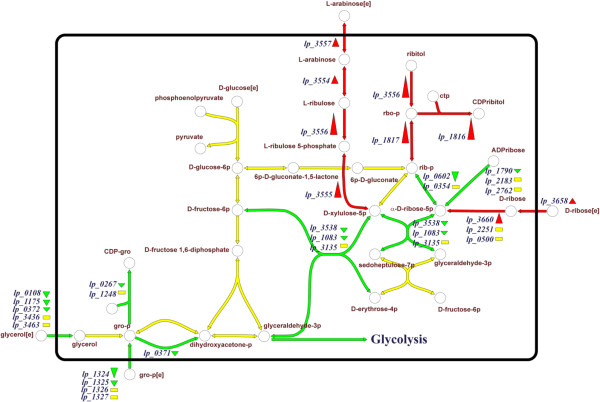
Through genome mining and mutagenesis we constructed derivatives that synthesize alternative WTA variants. The mutants were shown to completely lack WTA, or produce WTA and LTA that lack D-Ala substitution, or ribitol-backbone WTA instead of the wild-type glycerol-containing backbone. DNA micro-array experiments established that the tarIJKL gene cluster is required for the biosynthesis of this alternative WTA backbone, and suggest ribose and arabinose are precursors thereof. Increased tarIJKL expression was not observed in any of our previously performed DNA microarray experiments, nor in qRT-PCR analyses of L. plantarum grown on various carbon sources, leaving the natural conditions leading to WTA backbone alditol switching, if any, to be identified. Human embryonic kidney NF-κB reporter cells expressing Toll like receptor (TLR)-2/6 were exposed to purified WTAs and/or the TA mutants, indicating that WTA is not directly involved in TLR-2/6 signaling, but attenuates this signaling in a backbone independent manner, likely by affecting the release and exposure of immunomodulatory compounds such as LTA. Moreover, human dendritic cells did not secrete any cytokines when purified WTAs were applied, whereas they secreted drastically decreased levels of the pro-inflammatory cytokines IL-12p70 and TNF-α after stimulation with the WTA mutants as compared to the wild-type.
The study presented here correlates structural differences in WTA to their functional characteristics, thereby providing important information aiding to improve our understanding of molecular host-microbe interactions and probiotic functionality.
特定的植物乳杆菌菌株被作为有益健康的益生菌进行销售。细胞壁化合物(如壁磷壁酸和脂磷壁酸)在细菌生理学和益生菌-宿主相互作用中的作用和相互关系仍然不清楚。植物乳杆菌 WCFS1 具有切换 WTA 主链醛糖醇的遗传潜力,为在同基因背景下研究 WTA 主链修饰的影响提供了机会。
通过基因组挖掘和诱变,我们构建了合成替代 WTA 变体的衍生物。这些突变体被证明完全缺乏 WTA,或者产生缺乏 D-Ala 取代的 WTA 和 LTA,或者用核糖醇主链代替野生型甘油主链的 WTA。DNA 微阵列实验确定 tarIJKL 基因簇是合成这种替代 WTA 主链所必需的,并表明核糖和阿拉伯糖是其前体。在我们之前进行的任何 DNA 微阵列实验中,都没有观察到 tarIJKL 基因表达的增加,也没有在植物乳杆菌生长在各种碳源上的 qRT-PCR 分析中观察到增加,因此,如果存在导致 WTA 主链醛糖醇切换的自然条件,仍有待确定。表达 Toll 样受体 (TLR)-2/6 的人胚胎肾 NF-κB 报告细胞暴露于纯化的 WTA 和/或 TA 突变体,表明 WTA 不直接参与 TLR-2/6 信号传导,但以不依赖于主链的方式减弱这种信号传导,可能通过影响免疫调节化合物如 LTA 的释放和暴露来实现。此外,当应用纯化的 WTA 时,人树突状细胞不会分泌任何细胞因子,而在用 WTA 突变体刺激后,它们分泌的促炎细胞因子 IL-12p70 和 TNF-α 的水平明显降低,与野生型相比。
本研究将 WTA 的结构差异与其功能特征相关联,从而提供了有助于提高我们对分子宿主-微生物相互作用和益生菌功能理解的重要信息。