Department of Biochemistry and Metabolism, John Innes Centre, Norwich Research Park, Norwich NR4 7UH, UK.
The Sainsbury Laboratory, University of East Anglia, Norwich Research Park, Norwich NR4 7UH, UK.
Plant Cell. 2023 Sep 27;35(10):3809-3827. doi: 10.1093/plcell/koad204.
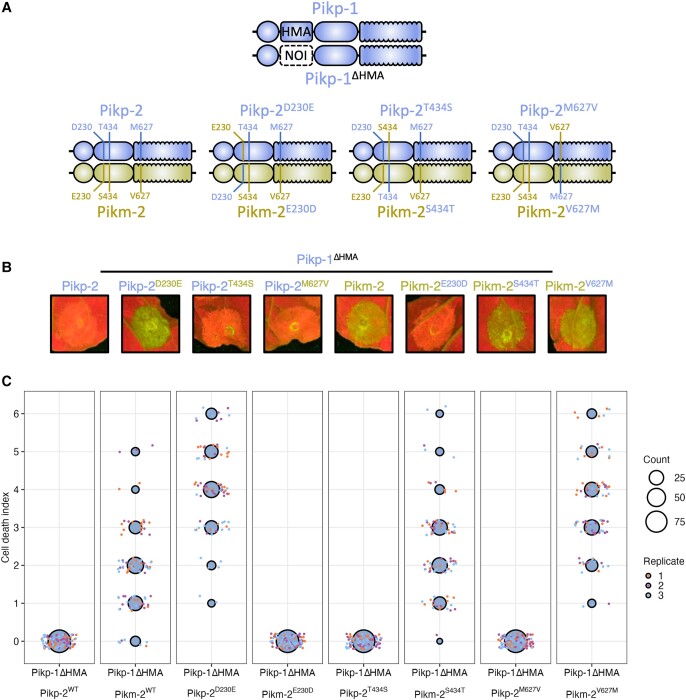
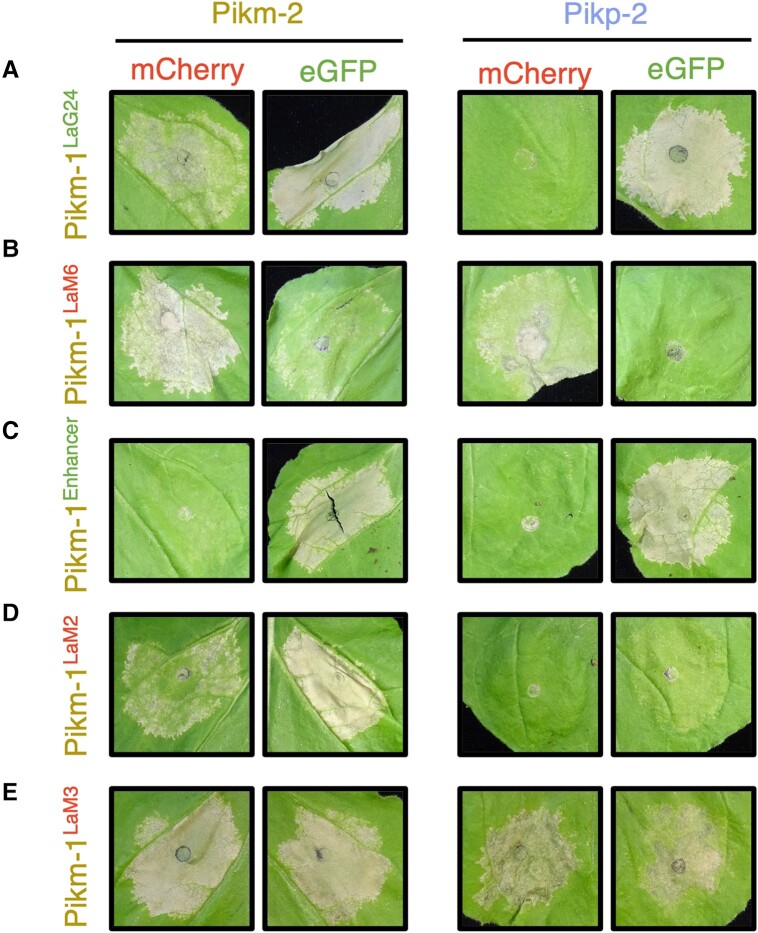
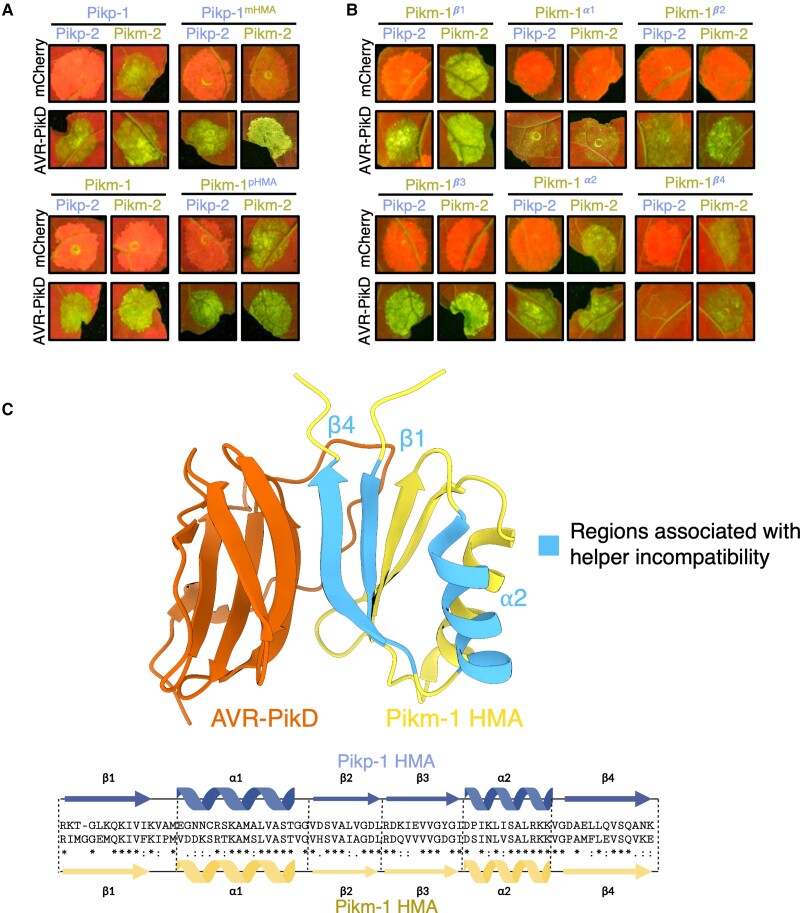
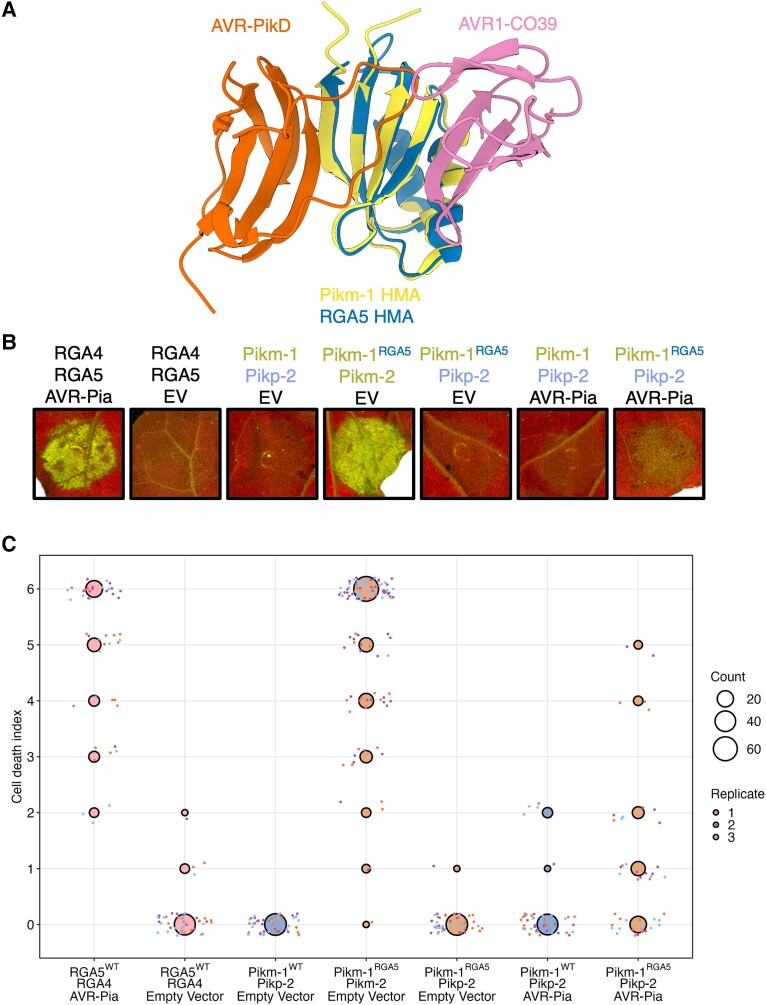
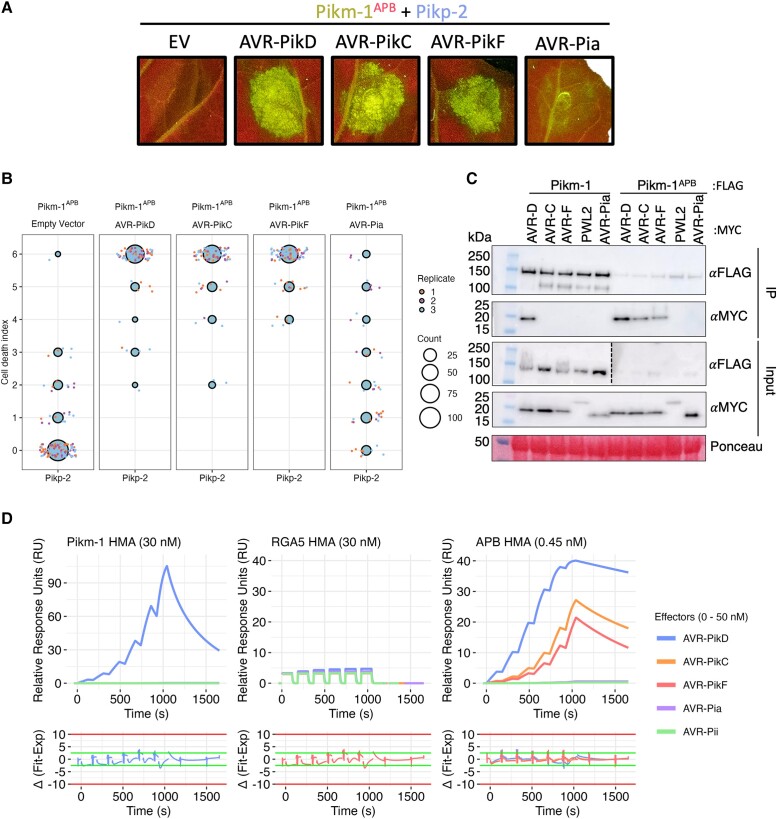
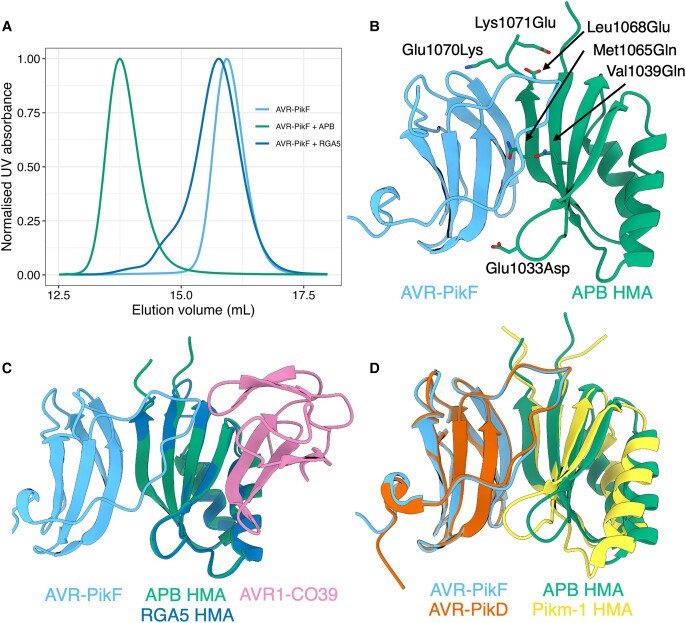
Engineering the plant immune system offers genetic solutions to mitigate crop diseases caused by diverse agriculturally significant pathogens and pests. Modification of intracellular plant immune receptors of the nucleotide-binding leucine-rich repeat (NLR) receptor superfamily for expanded recognition of pathogen virulence proteins (effectors) is a promising approach for engineering disease resistance. However, engineering can cause NLR autoactivation, resulting in constitutive defense responses that are deleterious to the plant. This may be due to plant NLRs associating in highly complex signaling networks that coevolve together, and changes through breeding or genetic modification can generate incompatible combinations, resulting in autoimmune phenotypes. The sensor and helper NLRs of the rice (Oryza sativa) NLR pair Pik have coevolved, and mismatching between noncoevolved alleles triggers constitutive activation and cell death. This limits the extent to which protein modifications can be used to engineer pathogen recognition and enhance disease resistance mediated by these NLRs. Here, we dissected incompatibility determinants in the Pik pair in Nicotiana benthamiana and found that heavy metal-associated (HMA) domains integrated in Pik-1 not only evolved to bind pathogen effectors but also likely coevolved with other NLR domains to maintain immune homeostasis. This explains why changes in integrated domains can lead to autoactivation. We then used this knowledge to facilitate engineering of new effector recognition specificities, overcoming initial autoimmune penalties. We show that by mismatching alleles of the rice sensor and helper NLRs Pik-1 and Pik-2, we can enable the integration of synthetic domains with novel and enhanced recognition specificities. Taken together, our results reveal a strategy for engineering NLRs, which has the potential to allow an expanded set of integrations and therefore new disease resistance specificities in plants.
利用工程改造植物免疫系统为克服由多种农业重要病原体和害虫引起的作物病害提供了遗传解决方案。修饰核苷酸结合富含亮氨酸重复(NLR)受体超家族的细胞内植物免疫受体,以扩大对病原体毒力蛋白(效应子)的识别,是工程抗病性的一种有前途的方法。然而,工程改造可能导致 NLR 自动激活,从而导致对植物有害的组成型防御反应。这可能是由于植物 NLR 以高度复杂的信号网络相关联,这些信号网络共同进化,通过繁殖或遗传修饰的变化会产生不兼容的组合,导致自身免疫表型。水稻(Oryza sativa)NLR 对 Pik 的传感器和辅助 NLR 共同进化,非共同进化等位基因之间的不匹配会触发组成型激活和细胞死亡。这限制了可以用于工程改造病原体识别并增强这些 NLR 介导的抗病性的蛋白修饰的程度。在这里,我们在 Nicotiana benthamiana 中剖析了 Pik 对中的不兼容性决定因素,发现 PIK-1 中整合的重金属相关(HMA)结构域不仅进化为与病原体效应子结合,而且可能与其他 NLR 结构域共同进化以维持免疫稳态。这解释了为什么整合域的变化会导致自动激活。然后,我们利用这些知识促进了新效应子识别特异性的工程改造,克服了最初的自身免疫惩罚。我们表明,通过错配水稻传感器和辅助 NLRs Pik-1 和 Pik-2 的等位基因,我们可以使合成结构域与具有新的和增强的识别特异性的结构域集成。总之,我们的结果揭示了一种工程改造 NLR 的策略,该策略有可能使植物中具有更大范围的整合,从而具有新的抗病特异性。